монтаж строительных лесов и их конструкция, установка навесных консолей и схема сборки, максимальная высота и другие размеры
Клиновые строительные леса – жесткая конструкция из металлических профилей, которую используют для проведения ремонтных или реконструктивных работ. Универсальное и удобное строение отлично подходит для монтажа различных конструкций или выполнения отделки и реставрации. Сегодня клиновые строительные леса считаются наиболее востребованными за счет повышенной безопасности и невысокой цены.
Особенности и элементы конструкции
Проведение строительно-монтажных или ремонтных работ требует использования лесов.
- подшивку кровли;
- утепление здания;
- отделку стен и фасадов;
- возведение загородных объектов.
Леса, выполненные из стальных труб и настила, способны выдержать нагрузку до 500 кг. Ключевая особенность подобных конструкций заключается в том, что собрать опорную опалубку можно при помощи соединения горизонтальных и вертикальных комплектующих. Крепление элементов между собой осуществляется посредством использования замкового механизма, который предотвращает неожиданное разъединение деталей.
Для сборки лесов монтажникам не приходится прикладывать значительные усилия, а эксплуатация конструкций не требует дополнительных вложений.
Вторая особенность клиновых лесов в том, что они безопасны для любых видов работ, связанных со строительством.

К преимуществам конструкций клинового типа относят ряд факторов.
- Высокая надежность конструкции. Соединение элементов клиновых лесов осуществляется с использованием замковых механизмов, что обеспечивает возможность для монтажа крупногабаритных грузов.
- Универсальность. Леса возводят для строительства и отделки разных объектов, вне зависимости от сложности проводимых работ и конфигурации сооружения.
- Простота монтажа и демонтажа конструкции. Ключевой инструмент – молоток для забивания и извлечения клина из фланцевых замков.
Полноценное использование клиновых лесов возможно только после их тщательной установки. При этом инженер и технический специалист должны проверить готовность конструкции и выписать разрешение на эксплуатацию сооружения.
Для устройства клиновых лесов используют следующие элементы:
- вертикальные металлические стойки;
- ригель для крепления горизонтальных стоек;
- горизонтальные стойки;
- башмаки, выступающие в качестве опорной части;
- стартовая деталь, представляющая собой вертикаль с фланцами;
- диагональ для связи элементов и обеспечения прочности конструкции;
- домкрат, выполненный в виде подпятника;
- лестница из металлических профилей;
- настил из дерева или металлического листа;
- кронштейны и другие крепежные элементы.
А также к составу конструкции относят стабилизирующую опору и навесную консоль.
Размеры
Клиновые леса востребованы на многих строительных площадках, где требуется организация фасадных работ. У подобных конструкций есть два типоразмера, которые определяют рабочую высоту сооружения.
- ЛСК-60. В этом случае максимальная высота лесов составит 60 м. Конструкции сооружают из труб, диаметр которых достигает 48 мм, а толщина стенок составляет 2 мм.
- ЛСК-100. Конструкции высотой 100 м собираются из труб диаметром 48 мм с толщиной стенок в 3 мм.
А также леса клинового типа способны образовать секции высотой по 2 м каждая. Ширина прохода между секциями не превышает 1-3 м.
Монтаж
Перед тем как приступить к монтажу конструкции, стоит проверить наличие необходимых инструментов, а также подготовить требуемое оборудование. Специалисту, который будет заниматься установкой конструкции, потребуется тщательно изучить схему ППР и повести проверку деталей на обнаружение поврежденных элементов.
Специалисту, который будет заниматься установкой конструкции, потребуется тщательно изучить схему ППР и повести проверку деталей на обнаружение поврежденных элементов.
Монтаж клиновых лесов выполняется с учетом требований строительных нормативов, к числу которых относятся СНиП 12-03-2001 и различные ГОСТы. А также необходимо принимать во внимание требования техники безопасности: выполнять работы в защитной одежде и с использованием надежных инструментов.
Монтаж выполняют в определенной последовательности.
- Сначала подготавливают площадку. Ее необходимо очистить от строительного мусора, снега, грязи, а также утрамбовать до получения ровной поверхности.
 Дополнительно потребуется вырыть канавы для стока воды.
Дополнительно потребуется вырыть канавы для стока воды. - Второй этап подразумевает установку лесов с углов здания. При этом важно, чтобы подъездные пути к объекту были организованы правильно, и конструкция не мешала проезду строительной техники.
- Далее под башмаки лесов устанавливают подкладки из дерева, толщина которых не превышает 45 мм в поперечном направлении. Иногда вместо башмаков устанавливают домкраты, которыми удается выровнять конструкцию с горизонтом.
- Четвертый этап – это монтаж стартовых элементов, крепление которых выполняют посредством продольных и поперечных связей.
- Когда предыдущие работы будут выполнены, мастер сможет приступить к основному этапу, который подразумевает установку вертикальных столбов в стартовые элементы. Крепление стоек выполняют, используя замковые механизмы или фланцы. Важно в процессе отслеживать уровень с помощью специального инструмента или отвесов.
- Чтобы обеспечить безопасность вертикальных стоек, конструкцию усиливают кронштейнами, прикрепляя ее к стене объекта. Дополнительно устанавливают навесные консоли.
- Затем на каждый ярус лесов укладывают настилы из дерева или металлических листов. А также возводят бортики, устанавливают лестницу для организации подъема и спуска.
- Следующий шаг заключается в усилении конструкции посредством монтажа диагонали, которую крепят с торца в двух прилегающих пролетах. Расстояние при этом не должно превышать 9-12 м.
- Далее мастер устанавливает перила, способные выдержать нагрузку от 70 кг. Особенно важно выполнить монтаж перил на верхних ярусах, чтобы предотвратить риск падения монтажников или штукатуров.
 Дополнительно потребуется вырыть канавы для стока воды.
Дополнительно потребуется вырыть канавы для стока воды.
Наконец, на последнем этапе выполняют заземление конструкции во избежание воздействия электрических зарядов во время грозы. После того как строительные работы будут выполнены, те же мастера выполняют демонтаж конструкции, придерживаясь обратного порядка. Сначала убирают диагональ, перила, настил, далее демонтируют лестницу и избавляются от мусора.
Сначала убирают диагональ, перила, настил, далее демонтируют лестницу и избавляются от мусора.
В процессе демонтажа мастера проводят рассортировку деталей с их последующей упаковкой.
Транспортировка и хранение
Недостаток клиновых лесов в том, что они не полностью покрыты краской. Некрашеные элементы требуют соблюдения особых условий для проведения надежной транспортировки и обеспечения качественного хранения. Перед этими мероприятиями требуется законсервировать комплектующие солидолом или другой смазкой. Перевозка оборудования осуществляется на специальном транспорте, способном вместить все составные части комплекта.Клиновые строительные леса – универсальная и безопасная конструкция, которая подходит для выполнения отделочных, ремонтных и реставрационных работ, а также для возведения различных зданий и сооружений.
Что такое строительные леса и их преимущества в строительно-монтажных работах
Нередко бывает так, что необходимые ремонтные или строительные работы требуется осуществлять на высоте. Приставные лестницы и стремянки не всегда являются спасением. В этом случае помогут строительные леса.
Определение строительных лесов
Данная конструкция напоминает переплетение металлических опор с элементами жесткости. В комплектации такого устройства входят настилы, зачастую, из дерева. Монтажные леса помогают при проведении строительных или ремонтных работ на высоте. На них могут разместиться строители и можно требуемый инструмент.
Монтажные леса помогают при проведении строительных или ремонтных работ на высоте. На них могут разместиться строители и можно требуемый инструмент.
Основные типы строительных лесов
Существуют такие разновидности лесов: рамные, клиновые, хомутовые.
Рамные леса
Это оборудование состоит из вертикальных (в некоторых случаях вертикальных) рам, соединенных между собой связями горизонтального и диагонального типа. Сборка осуществляется быстро, без применения специнструмента. В составе данных конструкций боковые рамы с интегрированной лестницей, горизонтальные и вертикальные связи, башмаки. Модели просты в установке, отличаются доступностью, поэтому они довольно популярны для ремонта и отделки прямых фасадов. Соединение элементов в единую систему происходит с помощью флажковых замков.
Главные характеристики:
- нормативная нагрузка – до 200 кг/м2;
- наибольшая высота для проведения кладочных работ – 25 м;
- наибольшая высота для проведения облицовки – 40 м.

Конструктивные особенности клиновых лесов
Фиксация элементов конструкции происходит с помощью клиньев из стали, вбиваемых молотком. Главные характеристики:
- нормативная нагрузка – не более 500 кг/м2;
- высота поднятия конструкции – около 120 м при диаметре труб 6 см;
- надежность.
Такие строительные леса часто используются в судо- и авиастроении, при установке сцен, реставрации фасадов, в кладочных работах.
Хомутовые конструкции
Они характеризуются довольно высокой сложностью сборки, компенсируемой достоинствами:
- конструкция вывести можно на 80 м;
- показатель нормативной нагрузки не превышает 250 кг/м2;
- возможность использования для работ на сложных фасадах, на большой площади.
Для сборки применяют глухие и поворотные хомуты.
Преимущества использования в строительно-монтажных работах строительных лесов
К сильным сторонам относят:
- повышенные показатели прочности и стойкости к нагрузкам;
- обширная рабочая зона на строительных лесах может разместиться даже 10 работников;
- универсальность – леса хорошо подходят для любых отделочных работ на любой высоте;
- простота установки и нет необходимости применять сложные инструменты и особые навыки;
- мобильность.

Особенности использования строительных лесов
Учитывая риски для сотрудников строительных и ремонтных бригад при использовании лесов, разработаны специальные правила, которые сводят к минимуму риск падения и прочих травм.
Если сравнивать с первыми модификациями, то отличие лесов новых поколений в большей устойчивости и продуманности конструкции, при этом наиболее популярны маловесные сооружения. Их установка и демонтаж не требует много времени, перевозка происходит с незначительными расходами, а использование возможно независимо от рельефа местности и конфигурации сооружения.
Преимущества сотрудничества с компанией Піонер
Компания Пионер ЮА является профессиональным оператором в сфере строительных лесов. Многолетний опыт производства и применение лесов на объектах разной сложности позволяют компании гарантировать покупателям надежность конструкций, функциональность и безопасность.
ПАСПОРТ СТРОИТЕЛЬНЫЕ ЛЕСА ЛСР 40
Леса строительные приставные рамные ЛРС 40
1. Назначение изделия
Назначение изделия
Леса рамные приставные ЛРС-40 предназначены для отделочных и ремонтных работ на фасадах зданий высотой до 40 метров, и нормативной поверхностной нагрузкой до 200 кг. На распределённый квадратный метр. Леса ЛРС-40 изготовлены в соответствии с ТУ 5225-001-38525343-2013.
2. Техническая характеристика ЛРС-40.
Леса ЛРС-40 изготовлены из стальной трубы диаметром 42мм с порошковой окраской. В строительных лесах ЛРС-40 сборка производится методом труба в трубу, используются рамы с вальцовкой, что обеспечивает жёсткость и надежность конструкции.
| Максимальная высота лесов | 40 метров |
| Высота яруса | 2 метра |
| Шаг яруса вдоль стены | 2 или 3 метра |
| Ширина яруса | 1 метр |
| Количество ярусов настила, не менее | 1 рабочий |
| Нормативная поверхностная нагрузка | 200 кг/м2 |
| Рама ЛРС- 40 изготовлена из круглой трубы | Ø 42 мм |
| Связь горизонтальная/диагональная | Труба Ø 25 мм |
| Размер между осей – горизонтальная связь | 3050 мм |
| Размер между осей – диагональной связи | 3292 мм |
| Ригель изготовлен из профильной трубы | 60х30 мм |
Примечание: При установке лесов ЛРС–40 в случаях, когда устанавливается узкая колонна строительных лесов, для обеспечения жесткости конструкции крепление к стене осуществлять в каждой секции конструкции и устанавливать диагонали в каждой ячейке.
Нагрузку при этом производить по спец. проекту ППР
3. Конструкция лесов ЛРС-40
Леса рамные приставные ЛРС-40 представляют собой пространственную каркасно-ярусную систему, смонтированную из трубчатых элементов: рам с лестницами, без лестниц, рам с торцевым ограждением, установленных в опорные башмаки или винтовые домкраты, соединённые между собой диагональными и горизонтальными связями, при помощи флажковых
фиксаторов. На опорную рамную конструкцию устанавливаются ригеля 2 м. или 3м. при фасадных работах. На ригеля укладываются деревянные щиты настилы, изготовленные из доски
20-25 мм. со специальным фиксирующим от опрокидывания замком. По желанию заказчика настилы обрабатываются огнеупорной пропиткой.
4. Элементы лесов и их описание
Рама без лестницы
Рама без лестницы ЛРС-40 изготовлена из электросварной трубы 42мм. Имеет габаритные размеры – высота – 2000 мм, ширина – 1000 мм. На раму приварены 4 флажковых фиксатора. Рама без лестницы является основным элементом и служит для наращивания лесов до необходимой высоты.
На раму приварены 4 флажковых фиксатора. Рама без лестницы является основным элементом и служит для наращивания лесов до необходимой высоты.
Рама с лестницей
Рама с лестницей ЛРС-40 изготовлена из трубы, 42 мм. Имеет габаритные размеры – высота – 2000 мм, ширина – 1000 мм. Является основным элементом и служит для наращивания лесов до необходимой высоты. На рамах предусмотрены замки с фиксатором для крепления диагональных стяжек и связей. Рама с лестницей является также межэтажным переходом для рабочих для их вертикального перемещения.
Горизонталь
Связь горизонтальная изготовлена из трубы 25 мм. Имеет габариты – длина 3050 мм, длина 2000мм. Служит для связи вертикальных рам и обеспечения необходимой дистанции между ними.
Диагональ диагональная стяжка
Связь диагональная изготовлена из трубы 25мм. Имеет габариты – длина 3292 мм, длина 2000 мм. Служит для обеспечения устойчивости и жесткости каркаса в горизонтальной плоскости, крепятся к рамам с помощью флажковых соединений.
Ригель
Ригель изготовлен из профильной трубы 60х30мм. Имеет габариты – длина 3065 мм, длина 2000 мм. Служит опорой для укладки деревянного настила. Ригели настилов навешиваются с помощью кронштейнов на верхние связи смежных рам на ярусах, предусмотренных под настилы. Сначала укладываются металлические ригеля, а затем укладывается деревянный настил.
Башмак опорный
Башмак опорный. Имеет габариты – 120*120 мм, O 40 мм, масса 0,9 кг Служит основанием нижнего ряда вертикальных рам лесов, принимает на себя вертикальные нагрузки от лесов и передает их на грунт посредством дополнительных деревянных подкладок.
Настил деревянный
Настил деревянный. Имеет габариты – ширина 1000 мм, длина 980 мм, масса 20 кг, изготавливается из досок 2-го сорта, расстояние м/у досками до 100мм, толщиной 20-25мм. Служит для горизонтального перемещения рабочих и принятия нагрузок, возникающих от бригад рабочих и строительно-отделочных материалов.
Анкерный кронштейн
Кронштейн – служит для крепления строительных лесов. Изготавливается из полосы 25*4 мм.
Крепление конструкции лесов к стене происходит с помощью кронштейнов и анкера, крайние стойки рам анкируют через 4 метра, внутренние стойки рам анкируют через 6 метров в шахматном порядке. Анкерные болты в комплекте с кронштейнами не поставляются.
Винтовой домкрат
Регулируемая опора – винтовой механизм, предназначенный для выравнивания вертикальных элементов строительных лесов. Необходим, при монтаже конструкций с небольшими неровностями опорной площадки. Комплектуется по письменной заявке заказчика строительных лесов.
5. Порядок сборки.
1. Монтаж и демонтаж лесов должен производиться под руководством ответственного производителя работ, который должен:
– изучить конструкцию лесов;
– составить схему установки лесов для конкретного объекта, согласно ППР;
– составить перечень потребных элементов;
– произвести согласно перечню приемку комплекта лесов со склада с отбраковкой
поврежденных элементов.
– знать и использовать всю нормативную документацию, связанную с техникой безопасности в строительстве СНиП 12 -03-2001.
2. Рабочие, монтирующие леса, предварительно должны быть ознакомлены с конструкцией и проинструктированы о порядке монтажа и способах крепления лесов к стене.
3. Леса должны монтироваться на спланированной и утрамбованной площадке, с которой должен быть предусмотрен отвод воды. Площадка под леса должна быть горизонтальной в продольном и поперечном направлениях. Она должна быть огорожена от возможного наезда строительной техники в зону установки фасадных лесов. Категорически запрещаетсяустанавливать леса на обледеневшее или неочищенное от снега основание.
4. Монтаж лесов следует производить, начиная от угла здания, соблюдая при этом последовательность установки отдельных элементов.
I Этап.
На подготовленной утрамбованной площадке установить деревянные подкладки толщиной не менее 40 мм, и башмаки согласно ППР проекту производства работ.
II Этап.
Установить на подпятники две смежные рамы первого яруса, соединить их горизонталями
и диагоналями связями в шахматном порядке.
Внимание!!! Вертикальные рамы следует устанавливать строго по уровню.
Аналогично монтируем следующие рамы строительных лесов до необходимой длинны.
III Этап.
Согласно ППР произвести крепление лесов к стене, при этом необходимо четко соблюдать вертикальность рам.
IV Этап.
Установить ригеля и деревянные настилы (металлические настилы) на первый рабочий ярус. При необходимости установить монтажный или дополнительный ярус. При укладке настила проверяются прочность закрепления и отсутствие возможности сдвигов. Зазоры между досками настила принимают не более 100 мм. Выступы досок за поверхность щита не должны превышать 30 мм. щиты настила могут быть соединены по длине внахлестку.
V Этап.
Установить рамы второго яруса, соединить их горизонтальными и диагональными связями, диагонали установить в шахматном порядке. В некоторых случаях для придания жесткости конструкции установка диагоналей осуществляется в каждой ячейке.
VI Этап.
Крепление (анкеровка) лесов к стене осуществить пробками или крюками с втулками посредством анкерных кронштейнов в шахматном порядке. Крайние стойки должны быть закреплены через 2 метра, внутренние через 4 метра, в шахматном порядке. В некоторых случаях для придания жесткости конструкции крепление лесов к стене осуществляется в каждой ячейке.
Повторяя эти этапы, набрать необходимую высоту лесов.
ВНИМАНИЕ !!!
Леса устанавливать четко по отвесу. Монтаж диагональных связей и крепление лесов к стене производить одновременно с монтажом лесов.
При установке лесов для каменной кладки наращивание их производится постепенно по мере возведения кладки стен здания. Укладку настилов и установку перил производить одновременно. В лесах для отделочных работ допускается укладывать щиты настила в шести ярусах, а для каменных только на двух верхних ярусах. Демонтаж лесов допускается лишь после уборки с настилов остатков материалов, инвентаря и инструментов. До начала демонтажа лесов производитель работ обязан осмотреть их и проинструктировать рабочих о последовательности и приемах разборки, а также о мерах, обеспечивающих безопасность работ. Демонтаж лесов следует начинать с верхнего яруса в последовательности, обратной последовательности монтажа. Демонтированные элементы перед перевозкой рассортировать. Крупногабаритные элементы связывать в пакеты. Произвольное сбрасывание элементов лесов категорически запрещается.
Укладку настилов и установку перил производить одновременно. В лесах для отделочных работ допускается укладывать щиты настила в шести ярусах, а для каменных только на двух верхних ярусах. Демонтаж лесов допускается лишь после уборки с настилов остатков материалов, инвентаря и инструментов. До начала демонтажа лесов производитель работ обязан осмотреть их и проинструктировать рабочих о последовательности и приемах разборки, а также о мерах, обеспечивающих безопасность работ. Демонтаж лесов следует начинать с верхнего яруса в последовательности, обратной последовательности монтажа. Демонтированные элементы перед перевозкой рассортировать. Крупногабаритные элементы связывать в пакеты. Произвольное сбрасывание элементов лесов категорически запрещается.
Для защиты от атмосферных электрических разрядов леса оборудуются заземлением, соединенным с рамой при помощи шины. Элементы заземления поставляются по отдельному заказу.
6 . Эксплуатация лесов
1. Металлические рамочные леса могут быть допущены в эксплуатацию только после окончания их монтажа но не ранее с дачи их по акту лицу, назначенному для приемки главным инженером строительства с участием работника по технике безопасности.
Металлические рамочные леса могут быть допущены в эксплуатацию только после окончания их монтажа но не ранее с дачи их по акту лицу, назначенному для приемки главным инженером строительства с участием работника по технике безопасности.
2. Осуществляется проверка при приемке установленных лесов в эксплуатацию следующих моментов:
– соответствие собранного каркаса монтажным схемам и правильность сборки узлов;
– правильность и надежность крепления лесов к стене;
– правильность и надежность опирания лесов на основание
– правильность установки молнии приемников и заземления лесов;
– обеспечение отвода воды от лесов.
Важно! Обратите особое внимание на вертикальность стоек и надежность крепления лесов к стене!
1. Состояние лесов должен ежедневно проверять перед началом смены производитель работ или мастер, руководящий работами.
2. Настилы и лестницы лесов рекомендовано систематически очищать от мусора, остатков материалов, снега, наледи, а зимой посыпать песком.
3. Нагрузки на настилы лесов в процессе их эксплуатации не могут превышать пределов, указанных в паспорте.
4. Важно соблюдать следующие правила при подаче материалов на леса башенным краном непосредственно к рабочим местам:
– чтобы избежать ударов грузом по лесам, следует сначала производить подъем груза и
передвижение крана, а затем поворот стрелы и плавное опускание груза;
– на лесах должен находиться сигнальщик, который регулирует подачу грузов подачей сигналов
крановщику.
Каркас стационарного подъемника, при подаче материалов на леса, должен крепиться к стене независимо от лесов.
7. Указание мер безопасности
Рамные строительные леса необходимо надежно закрепить к стене по всей высоте. Исключено произвольное снятие крепления лесов к стене.
– У настила лесов должна быть ровная поверхность.
– Подъем людей на леса и спуск с них должен осуществлять исключительно по лестницам.
– На лесах следует вывесить плакаты со схемами перемещения людей, размещения грузов, а также величин допускаемых нагрузок.
– Запрещается подача на леса груза, превышающего допустимый, проектом.
– Установка защитных устройств необходима во избежание повреждений стоек, расположенных у проездов.
– Линии электропередач, расположенные ближе 5 м от лесов необходимо снять или заключить в деревянные короба.
– Леса необходимо надежно заземлить и оборудовать грозозащитным устройством.
– Категорически запрещается доступ людей в зону ведения работ, не занятых на этих работах, при монтаже и демонтаже лесов.
– При монтаже полых коробчатых, трубчатых конструкций принимают меры против попадания и скопления в них воды.
– Леса, расположенные вблизи проезда транспортных средств, ограждают отбойными брусками с таким расчетом, чтобы они находились на расстоянии не ближе 0,6 м от габарита транспортного средства.
– При совпадении точек крепления с проемами в стене леса крепят к несущим конструкциям стенам, колоннам, перекрытиям с внутренней стороны здания при помощи различных приспособлений и устройств.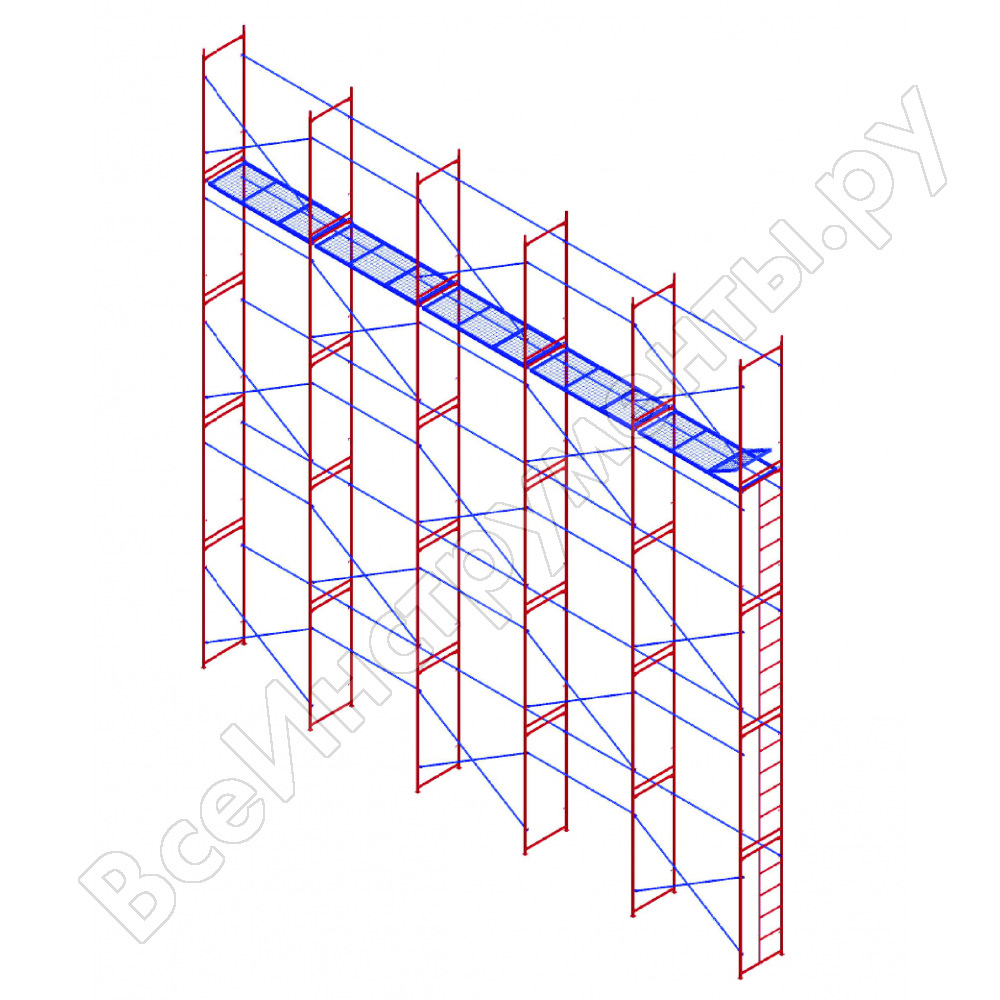 Не следует крепить леса к балконам, карнизам, парапетам.
Не следует крепить леса к балконам, карнизам, парапетам.
– Леса допускаются к эксплуатации после испытаний. При испытании лесов нормативной нагрузкой оцениваются их прочность и устойчивость, надежность настила и ограждений, заземления. Леса должны находиться под контрольной нагрузкой не менее 2 ч.
– Ввод лесов в эксплуатацию допускается после приемки комиссией, назначаемой руководителем строительной организации, и регистрируется в журнале учета по ГОСТ 26887-86. Если леса не использовались в течение месяца, то они допускаются к эксплуатации после приемки упомянутой комиссией. Леса следует эксплуатировать в соответствии с инструкцией предприятия-изготовителя и требованиями СНиП 12-03-2001.
– Рабочие должны быть ознакомлены с правилами работы на лесах, со схемами нагрузок, содержащими сведения о допустимых грузах и порядке их размещения.
Следует предусмотреть меры, чтобы обеспечить безопасный спуск людей с рабочего места при возникновении аварийной ситуации.
Кроме требований мер безопасности настоящего паспорта необходимо также выполнять требования СНиП 12-03-2001 Безопасность труда в строительстве и ПОТ РМ-012-2000 Межотраслевые правила по охране труда при работе на высоте.
8. Стандартная схема крепления лесов.
При сборке строительных лесов ЛРС-40 для придания жесткости конструкции, крепление лесов к стене осуществляется крайние рамы через 2 метра, внутренние через 4 метра в шахматном порядке, допускается крепление (анкеровка) в каждой ячейке и к каждой раме по всей площади лесов.
q = 200 кг/м2
Pp ≤ P доп., где Р доп. = 2600кгс.
9. Транспортирование и хранение
Транспортирование лесов может осуществляться транспортом любого вида в соответствии с действующими для данного вида транспорта правилами перевозки грузов).
1. Элементы лесов должны быть рассортированы по видам риге ль, стойка, связь и т.д. и связаны в пакеты проволокой диаметром не менее 4 мм в две нитки со скруткой не менее 2-х витков, перед транспортированием, при этом мелкие детали следует упаковать в ящики.
2. Сбрасывать элементы с транспортных средств при разгрузке запрещается.
3. Хранение лесов должно осуществляться по группе хранения ОЖ4 в соответствии с ГОСТ 15150-69.
4. В случае длительного хранения элементы лесов укладываются на подкладки, исключающие соприкосновение их с грунтом.
5. Металлические поверхности лесов, не имеющие лакокрасочных покрытий, при длительном хранении должны подвергаться консервации солидолом С по ГОСТ 4366 – 75 или другой равноценной смазкой.
Гарантии изготовителя.
ООО «ГРУППА ПРОФИ» гарантирует соответствие рамных строительных лесов ЛРС-40 требованиям настоящего паспорта при соблюдении потребителем условий эксплуатации, транспортирования и хранения.
Срок гарантии 12 месяцев со дня поступления потребителю.
Дата выпуска ________________________
Комплектовочная ведомость
| Наименование элемента | Количество, шт |
| Рама с лестницей | |
| Рама без лестницы | |
| Связь горизонтальная | |
| Связь диагональная | |
| Балка-ригель | |
| Опора | |
| Щит настила | |
| Анкерный кронштейн |
Внимание: количество кронштейнов, настилов и лестничных маршей, а также наличие
бортовых досок определяет заказчик строительных лесов согласно ППР для каждого объекта.
Монтаж и демонтаж строительных лесов – лучшие цены за м2
Мы предлагаем комплексную услугу по установке строительных лесов, которая включает в себя подготовку поверхности для монтажа строительных лесов, сборку элементов, закрепление конструкции на фасаде здания, установку строительной сетки и последующий демонтаж. Ниже представлены цены на монтаж и демонтаж лесов за метр квадратный.
Монтаж хомутовых лесов – от 180 р/м2
Монтаж клиновых лесов – от 120 р/м2
Монтаж рамных лесов – от 120 р/м2
Демонтаж лесов – от 80 р/м2
Монтаж и демонтаж фасадной сетки от 10 р/м2
У нас работают монтажники с большим опытом работы, что позволяет обеспечивать высокую скорость сборки строительных лесов. Как правило, монтаж и демонтаж лесов с нашей помощью в 2-3 раза быстрее самостоятельной сборки.
На стоимость монтажа влияет множество факторов, которые учитываются при детальном расчёте, окончательную стоимость монтажа уточняйте у наших специалистов по телефону +7 (495) 542-09-86.
Основные этапы монтажа и демонтажа строительных лесов
Если вы взяли леса фасадные в аренду, перед началом монтажа строительных лесов внимательно изучите их конструкцию, составьте схему установки, подготовьте перечень необходимых элементов конструкции. Монтажники, осуществляющие работы по сбору конструкции, должны быть проинструктированы о возможных способах крепления строительных лесов к стене.
Леса необходимо монтировать на специально отведенной для этих целей площадке. Грунт на площадке должен быть тщательно утрамбован, а поверхность площадки должна быть строго горизонтальной во всех направлениях. Монтаж строительных лесов осуществляется поярусно на всю длину объекта с использование подъемных механизмов.
1 этап
Установите деревянные подкладки и башмаки. Точки опор строительных лесов должны обязательно находиться в одной и той же горизонтальной плоскости.
2 этап
Установите две смежные рамы первого яруса в башмаки. Соедините рамы диагональными и горизонтальными связями. С шагом три метра установите смежные рамы. Рамы также необходимо соединить связями. Повторяйте данную манипуляцию необходимо количество раз для достижения необходимой длины участка строительных лесов. По краям требуется установить рамы ограждений.
3 этап
Установите следующие рамы второго яруса. Соедините рамы диагональными и горизонтальными связями. Диагональные связи необходимо устанавливать в шахматном порядке по отношению к друг другу. Для монтажа и укладки деревянных настилов используйте ригели.
4 этап
Установите наклонные лестницы, необходимые для подъема и спуска строительного персонала. Лестницы устанавливаются в люках, специально отведенных для данной цели.
5 этап
Строительные леса крепятся к поверхности стены специальными крюками с втулками через кронштейны или хомуты. Крепление осуществляется в шахматном порядке через каждые 3-4 метра.
6 этап
Повторяя предыдущие этапы (3,4 и 5) можно набрать требуемую высоту строительных лесов.
7 этап
Необходимо установить на рабочих и предохранительных ярусах связи ограждения (торцевые и продольные). В местах подъема и спуска рабочего персонала на рабочий ярус обязательно установите продольные связи ограждений. Последний (верхний) ярус монтируется из стоек и рам ограждения высотой 1,5 метра.
Обратите внимание, процесс закрепления лесов к поверхности стены осуществляется одновременно с монтажом строительных лесов. Настилы укладываются одновременно с установкой связей ограждений.
Демонтаж строительных лесов
Демонтаж лесов возможен только после уборки с настилов остатков строительных материалов и инструментов. Демонтаж лесов необходимо начинать с последнего (верхнего) яруса в обратной последовательности. Перед транспортировкой демонтируемые элементы конструкции необходимо пересчитать и рассортировать. Габаритные элементы конструкции необходимо упаковать в пачки, малогабаритные элементы в ящики.
Хомутовые строительные леса ЛСПХ
Ремонт и строительство зданий сложной конфигурации требует применения особых оградительных конструкций. Благодаря использованию хомутовых лесов, это задача решается наилучшим способом.
Строение хомутовых лесов
Рамная конструкция из вертикальных, горизонтальных и диагональных элементов. Крепежом для соединения элементов рамы между собой выступают хомуты двух видов: жесткие и поворотные.
Вертикальной стойкой служит труба из 2-х или 3-миллиметровой стали сечением 48 мм. По длине различаются трубы 2-х и 4-метровые. Основание стоек служат опорные башмаки. В первом ярусе разнодлинные стойки чередуют. Для второго и всех последующих рядов используются только трубы 4 м. на последнем ярусе опять применяется принцип чередования. Соединение происходит «труба в трубу».
Горизонтальные и диагональные связи крепятся к вертикальным стойкам при помощи хомутов. Это позволяет изменять угол, высоту яруса и величину шага между стойками. Именно благодаря болтовому способу крепления, хомутовые леса применяют для работы со сложной формой фасада.
На продольные связи монтируется щитовой настил. Анкерным соединением хомутовые леса крепятся к фасаду здания.
Виды работ с хомутовыми лесами
Благодаря прочности и надежности (до 300 кг/м2) хомутовые строительные леса используют на различных строительных, ремонтных и монтажных работ. Их применение обосновано для возведения сцен, подиумов, навесов. Подвижный хомут позволяет состыковывать данный вид со строительными лесами других типов:
- Рамные;
- Штыревые;
- Клиновые.
Максимальная высота для проведения реконструкций фасадов – 40 м, для ведения кирпичной кладки – 20 м.
Для обеспечения безопасности рабочих хомутовые леса снабжены заземлением, страховочными бортиками, строительной сеткой.
Продажа и аренда — хомутовые строительные леса
Наша компания занимается продажей и арендой приставных строительных конструкций для проведения высотных работ. Хомутовые леса соответствуют ГОСТу 27321-87.
Ассортимент представлен новыми и б/у конструкциями. Стоимость хомутовых лесов несколько выше в сравнении с конструкциями других типов. Но они часто применяются там, где использование рамных или клиновых лесов невозможно: для обхода балконов, козырьков, при строительстве и ремонте куполов, водонапорных башен и других сооружений.
Монтаж и демонтаж хомутовых лесов несколько сложнее и требует большего времени. К установке допускаются рабочие, прошедшие инструктаж. сборка происходит под руководством и контролем специалиста. Поэтому мы предлагаем также услуги по монтажу и демонтажу строительных лесов.
Все вопросы по продаже и аренде строительного оборудования вы можете обсудить с нашим менеджером. Мы гарантируем поставку лесов в оговоренные сроки.
Сборка и строительные леса
Сборка и строительные леса
1Я не могу найти четкого определения, которое отличает сборку последовательностей от строительных лесов.
Насколько я понимаю,
Сборка = объединение считываний в контиги
Строительные леса = объединение контигов в каркасы (например, с использованием считывания парных концов)
Звучит правильно? Кажется, что за сборкой должны следовать строительные леса, но определения сборки даже не говорят о строительных лесах. Сможете ли вы собрать целый геном с помощью всего лишь «сборки»?
сборка последовательность действий • 4.8k просмотров • ссылка обновлено 5,7 года назад пользователем новичок & starf; 1.0k • написано 5,7 лет назад пользователем сарданьо & utrif; 10Контиги – это последовательности перекрывающихся ( контигов ous) считываний.Чтения с парным концом (или парой сопряжения) можно использовать для определения разрыва между двумя контигами. Зная промежуток, вы можете создать каркас, состоящий только из двух контигов, где N представляют промежуток между ними.
РЕДАКТИРОВАТЬ: Можете ли вы собрать весь геном просто “сборкой”?
Да. Строительные леса в любом случае не дадут вам больше информации о реальных базах; он просто пытается сказать вам, как упорядочены ваши контиги.
• ссылка обновлено 3.2 года назад, автор: Баран 35 тыс. • написано 5,7 лет назад пользователем новичок & starf; 1.0kАвторизуйтесь, прежде чем добавлять свой ответ.
Создание каркаса для сборки последовательности генома с использованием ABySS
Создание каркаса для сборки последовательности генома с использованием ABySSШон Д. Джекман, Энтони Г. Раймонд и Инанс Бироль
Канадский центр геномных наук Майкла Смита
Разработка библиотек для секвенирования генома на больших расстояниях, известных как библиотеки мат-пар или прыжков, позволяет контигам de novo сборка последовательности генома для сборки в каркасы, которые определяют порядок и ориентация этих контигов.Мы разработали для Программа для сборки последовательности генома de novo ABySS серия эвристические алгоритмы, каждый из которых определяет небольшой подграф граф каркаса, соответствующий определенной топологии и применяющий преобразование в этот подграф, чтобы упростить граф каркаса. Эти алгоритмы устраняют неоднозначности в графе каркаса и определяют контиги, которые могут быть собраны в каркас.
Современная технология короткого чтения генома дает миллиарды короткое чтение, каждые 150 пар оснований в длину, хотя эта длина постоянно увеличивается.Когда доступна эталонная последовательность генома, эти прочтения могут быть согласованы с этой ссылкой для определения одного варианты нуклеотидов, другие варианты малых последовательностей и большие структурная вариация. Когда эталонная последовательность недоступна, или когда эксперимент не хочет быть предвзятым ссылкой последовательность, последовательность чтения должна быть собрана de novo. Из-за пробелов в секвенирование и повторяющаяся последовательность генома, такая сборка часто фрагментируется на множество последовательностей, называемых контигами. Относительная порядок и ориентация этих контигов неизвестны, и проблема упорядочивания и ориентации этих контигов называется лесами.
Создание лесов осуществляется с использованием секвенирования парных концов, где оба концы одного и того же фрагмента ДНК секвенированы. Успех скаффолдинг ограничен размером библиотеки фрагментов и ее способность охватывать самые большие повторяющиеся последовательности генома. До недавно библиотека парного секвенирования была ограничена примерно 800 пар оснований. Новые технологии в построении библиотек позволили для создания библиотек мат-пар размером до 5000 п.н. Новый технологии секвенирования требуют разработки новых алгоритмов для полностью использовать технологию.Дальнейшее развитие нашей работы над Программное обеспечение для сборки секвенирования novo, ABySS, мы разработали новый алгоритм каркаса, способный создавать каркасы очень больших геномов. Мы использовали ABySS, чтобы сформировать сборку последовательности генома белых ель, Picea glauca, геном которой 20 гигабаз составляет примерно семь раз больше человеческого генома.
Каркасный граф состоит из вершин, представляющих последовательности и ребра, представляющие набор парных чтений, которые указывают порядок и ориентация этих двух контигов, а также приблизительная оценка расстояния, разделяющего две последовательности.Наши строительные леса алгоритм реализован в виде серии эвристических графов преобразования, каждое из которых определяет небольшой подграф, соответствующий топология, типичная для определенного геномного признака, например, повтор последовательность, и применяет преобразование к этому подграфу, чтобы последовательно упростить граф каркаса. Эти алгоритмы устраняют неоднозначности в графе каркаса и идентифицируют контиги, которые собран в подмости.
Мы сравниваем наши результаты строительных лесов с результатами сборки de novo программные пакеты ALLPATHS-LG, SGA и SOAPdenovo.Эти три программных пакета и ABySS были используется для сбора данных короткочитаемого секвенирования образца человека NA12878. Подмости этих сборок были выровнены по эталонный геном человека для определения смежности и правильности строительных лесов и сравните производительность этих инструментов.
Терминология
Каркасный граф – это ориентированный граф, в котором каждая вершина представляет собой последовательность контигов. Каждое направленное ребро e = (u, v) представляет собой пучок парные чтения, объединяющие два контига, где для каждой пары одно чтение выравнивается по последовательности вершины u, а другое чтение выравнивается по последовательность вершины v.Направление кромки (u, v) указывает на то, что ориентация считываний показывает, что contig v появляется после contig u в геноме.
Каркасный граф – это кососимметричный граф. Каждая вершина u имеет дополнительная вершина ~ u, последовательность которой является обратным дополнением к последовательность вершины u. Каждое ребро (u, v) имеет дополнительное ребро (~ v, ~ u). Любые операции манипулирования графом поддерживают это свойство.
Свойства связаны с каждой вершиной, например, длина последовательность и каждое ребро, например количество парных чтений, поддерживающих край.
Обозначение
- g – ориентированный граф.
- u, v и w – вершины.
- e = (u, v) – направленное ребро.
- g [u] .x – некоторое свойство x вершины u.
- g [e] .x – некоторое свойство x ребра e.
- g [u] .outdeg – внешняя степень вершины u.
- g [u] .indeg – внутренняя степень вершины u.
- g [u] .l – длина последовательности вершины u.
- g [e] .n – количество парных чтений, поддерживающих край e.
Свойство g [e] .n, где e = (u, v), – это количество парных чтений. где одно чтение отображается в последовательность вершины u, а второе чтение отображается в последовательность вершины v с ориентациями, которые согласуются с направление кромки e.
Процедуры манипуляции с графиком
Следующие процедуры манипулирования графами используются для определения алгоритмы.
- remove_vertex (вершина u, граф g): удалить вершину u и ее инцидентные ребра из графа g.Дополнительная вершина ~ u и его инцидентные ребра также удаляются из графа g.
- add_edge (вершина u, вершина v, граф g): добавить ребро (u, v) и его дополнение (~ v, ~ u) к графу g.
- remove_edge (edge e, graph g): удалить ребро e и его дополнительное ребро ~ e графа g.
Алгоритм scaffolding реализован в виде последовательности эвристик. что каждый идентифицирует подграфы, соответствующие определенной топологии, а затем преобразовать этот подграф каким-либо образом, добавляя и удаляя вершины и края.Хотя это не показано явно в приведенных ниже алгоритмах, для каждый алгоритм – это модификации графа, определяемые этим алгоритмы выполняются одновременно, так что результат не зависят от порядка, в котором вершины и ребра посещаются обход графа.
Фильтр графика
Короткие контиги и ребра, которые плохо поддерживаются парным чтением, являются удален с графика. Вершины короче s bp удаляются из граф, и ребра, поддерживаемые менее чем n парными чтениями, являются удалены с графика, где s и n – параметры алгоритма.Короткие контиги могут повторяться, и в результате могут возникнуть проблемы с опорой на края. из чтений, которые неправильно сопоставлены сборке.
процедура filter_graph (график g):
для вершины u в g
если g [u] .l Вилка Resolve
Вилка состоит из двух ребер (u, v1) и (u, v2) без ребра (v1, v2) или (v2, v1) присутствует в графе g. Вершины v1 и v2 известны следовать за u, но неизвестно, следует ли v1 за v2 или наоборот.Там может существовать ребро в исходном графе g0 до фильтрации, которое разрешает эту неоднозначность относительного порядка v1 и v2. Мы проверяем существуют ли ребра (v1, v2) и (v2, v1) в графе g0, и если существует ровно одно такое ребро, мы добавляем его в граф g.
процедура resolve_forks (график g0, график g):
для ребер (u, v1), (u, v2) в g
если существует (v1, v2) в g0 и не существует (v2, v1) в g0
add_edge (v1, v2, g)
если существует (v2, v1) в g0 и не существует (v1, v2) в g0
add_edge (v2, v1, g)
Рисунок 1.Вилка ResolveКончики чернослива
Кончик - это вершина, которая образует короткую шпору, отходящую от другого непрерывный путь. У него внутренняя степень = 1 и исходящая степень = 0. Эти подсказки удаляются с графика.
процедура prune_tips (график g):
для ребра (u, v) в g
если g [u] .outdeg> 1 и g [v] .indeg = 1 и g [v] .outdeg = 0
remove_vertex (v, g)
Рис. 2. Кончики обрезкиУдалить повторы
Повтор - это последовательность, которая многократно встречается в геноме, собран.Повторение, достаточно маленькое, чтобы его можно было охватить парными чтениями вызывает контрольные транзитивные ребра в графе каркаса. Эти транзитивные ребра в конечном итоге будут удалены из каркаса, но удаление этих транзитивных ребер из графа каркаса удаляет та самая информация, которая разрешила бы повторение. Перед удалением переходные ребра, мы идентифицируем и удаляем вершины, образованные повторяющаяся последовательность.
процедура remove_repeats (график g):
для ребер (u1, w1), (u2, w2), (u1, v), (v, w1), (u2, v), (v, w2) в g
remove_vertex (v, g)
Рисунок 3.Удалить повторыУдалить переходные края
Ребро (u, w) транзитивно, если в графе существует путь из вершина u в вершину w. Поиск и удаление всех транзитивных ребер из График - потенциально трудоемкая операция. Однако мы можем легко найти переходные ребра с максимальной длиной пути, равной двум, которые удален с графика.
процедура remove_transitive_edges (график g):
для ребер (u, v), (v, w), e = (u, w) в g
remove_edge (e, g)
Рисунок 4.Удалить переходные краяУдалить закрытые пузыри
Пузырь на графике Хэн Ли определил следующим образом:
Пузырь - это направленный ациклический подграф с одним источником и одиночный сток, имеющий как минимум два пути между источником и раковина. Закрытый пузырь - это пузырь без входящих краев из или выходящие ребра в другие части всего графа, кроме исток и сток вершины. Закрытый пузырь прост, если есть ровно два пути между источником и стоком; иначе это сложно.
Отсутствует край в графе каркаса из-за отсутствия покрытия, обычно приводит к простому замкнутому пузырю, где две вершины, v1 и v2, как известно, встречаются между двумя другими вершинами, u и w, но отсутствие ребра между v1 и v2 означает, что порядок v1 и v2 равен неизвестный. Эта ситуация разрешается удалением вершин v1 и v2 из графа и добавив ребро (u, w).
процедура remove_simple_bubbles (график g):
для ребер (u, v1), (v1, w), (u, v2), (v2, w) в g
remove_vertex (v1, g)
remove_vertex (v2, g)
add_edge (u, w, g)
Рисунок 5.Удаление простых пузырьковСложные пузырьки можно идентифицировать и удалять аналогичным образом. Топологический упорядочение графа вычисляется с использованием поиска в глубину (DFS). Обратные ребра, возникающие в результате циклов в графе, безопасно игнорируются, поскольку пузыри по этому определению являются ациклическими подграфами. В топологический порядок, вычисленный DFS, группирует вершины замкнутый пузырь вместе в топологическом порядке, и это свойство используется для выявления и удаления сложных закрытых пузырьков из скаффолд граф.
Удалить слабые кромки
Ребро e = (u1, v2) определяется как слабое, если существуют ребра e1 =
(u1, v1) и e2 = (u2, v2) в графе, где g [e] .n Ребро (u, v) неоднозначно, если либо исходная степень u, либо
in-степень v больше единицы. Эти неоднозначные края удалены
из графика. Остальные кромки образуют непрерывные дорожки, которые собираются в
создать финальные строительные леса.Последовательности вершин пути
соединяются, перемежаются с пробелами, представленными серией
символ N, длина которого соответствует оценке расстояния
между этими двумя контигами. Оценщик максимального правдоподобия используется для
оцените расстояние между двумя контигами по выравниванию
парное чтение контигов. Возможно, что расстояние
оценка отрицательная, что указывает на то, что два контига на самом деле должны
перекрывать. Если такое перекрытие действительно обнаружено в перекрытии контигов
На графике два контига объединяются. Мы собрали данные секвенирования парных концов 101 пар оснований человека.
образец NA12878 с использованием ABySS. Эшафоты были
разделить на N, чтобы получить контиги. Контиги были привязаны к человеческому
ссылка GRCh47 с использованием BWA-SW с командной строкой Положение контигов в каждом каркасе сравнивали с
положение контигов, выровненных по эталонному геному, для идентификации
контиги, которые не совпадают в соответствии с их
расположение в эшафоте. Для последовательности контигов в каркасе
начальные позиции этих контигов в эталонном геноме должны быть
монотонно возрастающая последовательность или убывающая, если каркас
с обратным дополнением относительно ссылки, и аналогично
конечные положения также должны быть монотонно возрастающей последовательностью.В
каркас сломан на любой паре контигов, не удовлетворяющих этим требованиям.
критериев, и рассчитывается выровненный каркас N50. Этот анализ был повторен для сборок NA12878
ALLPATHS-LG, fermi, SGA и
SOAPdenovo. Сборка ALLPATHS-LG скачивается с
NCBI, а сборки fermi, SGA и SOAPdenovo были предоставлены
Хэн Ли, Джаред Симпсон и Руибанг Луо соответственно (см. Материалы).
Сборки ABySS, fermi, SGA и SOAPdenovo используют идентичные данные.
наборы. Сборка ALLPATHS-LG использует более глубокий 100-кратный набор данных и два
библиотеки сопряженных пар.Собранные ферми контиги были собраны
в каркасы с использованием алгоритма каркасов ABySS. Построение выровненного контига N50 в зависимости от количества точек останова
дает график, где лучшие сборки находятся в верхнем левом углу
угол.Построение выровненного каркаса N50 в зависимости от количества
контрольные точки каркаса дают аналогичный график, показывающий производительность
алгоритм строительных лесов. Данные секвенирования человеческого образца NA12878 (SRS000090) составляют 101 п.н.
Иллюмина читает. Было использовано восемь полос парных данных, четыре полосы парных
библиотека Solexa – 18483 и четыре переулка библиотеки Solexa – 18484:
ftp: // hengli: reichdata @ ftp.broadinstitute.org/NA12878-hs37d5-aln/NA12878-hs37.bam Четыре дорожки библиотеки пар матов Solexa – 30807 (SRX027699) были
использовал:
http://sra.dnanexus.com/experiments/SRX027699 Дж. Т. Симпсон, К. Вонг, С. Д. Джекман, Дж. Э. Шейн, С. Дж. М. Джонс, İ Birol
(2009).ABySS: параллельный ассемблер для данных короткой последовательности чтения.
Геномные исследования, 19 (6), 1117–1123. Gnerre S, MacCallum I, Przybylski D, Ribeiro F, Burton J, Walker B,
Шарп Т., Холл G, Ши Т., Сайкс С., Берлин А., Эйрд Д., Костелло М., Даза
Р., Уильямс Л., Николь Р., Гнирке А., Нусбаум С., Лендер Е. С., Джаффе Д. Б.
(2011).
Качественные черновые сборки геномов млекопитающих из массовых
данные параллельной последовательности.
Труды Национальной академии наук, 108 (4), 1513–1518. Ли Х. и Дурбин Р.(2010).
Быстрое и точное выравнивание с длинным считыванием с помощью преобразования Барроуза-Уиллера.
Биоинформатика, Epub. Марк А. ДеПристо, Эрик Бэнкс, Райан Поплин, Киран В. Гаримелла, Джаред Р.
Магуайр, Кристофер Хартл, Энтони Филиппакис, Гильермо дель
Ангел, Мануэль А. Ривас, Мэтт Ханна, Аарон МакКенна, Тим Дж. Феннелл,
Андрей Керницкий, Андрей Сиваченко, Кристиан Цибульскис, Стейси Б.
Габриэль, Дэвид Альтшулер и Марк Джей Дэйли
(2011).
Основа для обнаружения вариаций и генотипирования с использованием
данные секвенирования ДНК следующего поколения.Nature Genetics 43 (5), 491–498. Хэн Ли
(2012).
Изучение SNP на одном образце и вызов INDEL с помощью полногеномного
новая сборка.
Биоинформатика, предварительный доступ. Джаред Т. Симпсон и Ричард Дурбин
(2012).
Эффективная сборка больших геномов de novo с использованием сжатых данных
конструкции.
Исследование генома, 22 (3), 549–556. Руйцян Ли, Хунмей Чжу, Цзюэ Руань, Вубинь Цянь, Сяодун Фанг,
Чжунбинь Ши, Инжуй Ли, Шэнтин Ли, Гао Шань, Карстен Кристиансен,
Сонган Ли, Хуанмин Ян, Цзянь Ван и Цзюнь Ван
(2010).Сборка de novo геномов человека с массовым параллельным коротким чтением
последовательность действий.
Исследование генома, 20 (2), 265–272. Скомпилируйте арки, ссылки и установите tigmint в соответствии с инструкциями по компиляции арков и ссылок. Если вы работаете в нашей лаборатории, вам не нужно этого делать - каталог открыт для общего доступа. Перед использованием имеющихся у нас данных 10-кратного связанного чтения нам необходимо установить программу longranger от 10X genomics.Это довольно простой процесс - вам просто нужно перейти на веб-сайт longranger, взять ссылку и загрузить этот файл в подходящее место (в нашем случае - общий каталог Установить рабочий каталог. Здесь мы работаем с Diglossa , поэтому: Теперь мы запустим longranger, чтобы выполнить базовую обработку данных 10-кратного связанного чтения.Идите вперед и создайте каталог в 10x-diglossa-scaffold для хранения данных: Подготовьте сценарий После этого нам нужно сгенерировать файл кратностей штрих-кода. Мы можем сделать это с помощью сценария perl из пакета arks.Перед запуском этого сценария perl вам необходимо создать файл конфигурации, содержащий путь к обработанным связанным данным чтения сверху. В нашем каталоге Например, мой И моя общая структура каталогов будет выглядеть так: Теперь, когда мы создали этот файл имен файлов (FOFN или fofn), мы можем вычислить кратности штрих-кода. Это занимает около 30 минут для чтения файла размером 50 ГБ: Перед тем, как строить скаффолд, нам нужно загрузить сборку contig files / pacbio, которую мы хотим скаффолд: Итак, теперь наша структура каталогов выглядит примерно так: Наконец, мы готовы запускать арки. В вашем рабочем каталоге создайте последний каталог для хранения выходных данных arks: arks в основном запускается через make-файл, пример которого доступен на странице arks на github. Я уже частично отредактировал этот make-файл, чтобы упростить запуск arks, учитывая тот способ, которым мы его установили.Вы можете скачать мою отредактированную версию здесь. Мы можем увидеть параметры, которые принимает make-файл, загрузив файл и запустив его с помощью команды Вообще говоря, есть некоторые значения в make-файле, которые мы хотим изменить, в частности, параметры количества потоков для arks также плохо справляется с путями при его вызове, поэтому в Теперь, когда мы отредактировали make-файл Сборка Некоторые классы бактериальных вирусов и вирусов животных требуют временного присутствия молекул, известных как каркасные белки, которые необходимы для сборки предшественника прокапсида.Чтобы собрать прокапсид надлежащего размера, каждая субъединица вирусной оболочки должна принять правильную квазиэквивалентную конформацию из нескольких возможных вариантов, в зависимости от номера T капсида. В отсутствие каркасного белка белки оболочки вируса образуют капсиды неправильной формы и неправильного размера, которые не могут упаковать ДНК. Хотя каркасные белки не образуют икосаэдрических ядер внутри прокапсидов, икосаэдрически упорядоченное взаимодействие покрытие / каркас может объяснить, как каркас может вызывать конформационные различия между субъединицами оболочки.Чтобы идентифицировать сайты взаимодействия каркасного белка с решеткой белка оболочки бактериофага Р22, мы определили электронную криомикроскопию структуры прокапсидов, содержащих каркас и лишенных каркаса. Полученные в результате карты различий предполагают специфические взаимодействия каркасного белка только с четырьмя из семи квазиэквивалентных конформаций белка оболочки в решетке прокапсида T = 7 P22, подтверждая идею о том, что конформационное переключение субъединицы оболочки регулируется типом взаимодействий, которые она подвергается каркасному белку.Основываясь на этих результатах, мы предлагаем модель сборки прокапсида P22, которая включает в себя чередующиеся этапы, в которых субъединицы сначала покрывают, а затем скаффолдинги образуют самовзаимодействия, которые способствуют добавлению другого белка. Вместе покрытие и каркас обеспечивают перекрывающиеся наборы связывающих взаимодействий, которые управляют образованием прокапсида. Рекомендуемые статьиЦитирующие статьи (0) Copyright © 1999 The Biophysical Society. Опубликовано Elsevier Inc. Все права защищены. Сообщение Линь-Син Чена, Картика Анантарамана, Алона Шайбера, А. Мурата Эрена и Джиллиан Ф. Банфилд. С любыми вопросами о методах, описанных ниже, свяжитесь с нами по электронной почте ([email protected]). Введение Цель этого поста - описать пошаговую процедуру расширения каркаса и закрытия разрыва для значительного улучшения качества геномов, собранных в метагеноме (MAG).Эти стратегии описаны в нашей недавней статье под названием «Точные и полные геномы из метагеномов» (сокращенно CGM), которая доступна по адресу https://genome.cshlp.org/content/early/2020/03/18/ gr.258640.119.full.pdf + html. Пожалуйста, процитируйте эту публикацию, если вы будете следовать описанным здесь процедурам для анализа. В документе CGM обобщается история метагеномики с разрешением генома с примерами из публикаций, чтобы показать необходимость объединения геномов и ручного курирования MAG, чтобы избежать вводящих в заблуждение выводов.В нашей статье представлена пошаговая процедура создания черновика MAG, сгенерированного ручными или автоматическими инструментами объединения, для предотвращения ошибок (например, неправильно построенных строительных лесов, локальных ошибок сборки), закрытия зазоров в строительных лесах и, таким образом, создания высококачественных или полных MAG. (CMAG). Здесь мы подробно рассмотрим две основные стратегии, которые мы обычно используем для полного улучшения MAG: расширение каркаса и закрытие пробелов. Пристройка подмостей Удлинение каркаса использует считывания парных концов, картированных на данный каркас для удлинения его концов, (1) для получения полной последовательности гена, кодирующего белок или гена рРНК на концах, (2) для сборки его с другим каркасом на основе на перекрывающихся последовательностях в конце (3) для получения полного генома бактерий, архей, вирусов, фагов и т. д.В документе CGM говорится: «Строительные леса внутри бункера, которые не перекрываются в начале курирования, могут быть соединены после одного или нескольких раундов наращивания лесов. Этот процесс расширения, соединения и повторного отображения может продолжаться до тех пор, пока все фрагменты не составят единую кольцевую последовательность. Следует отметить, что чтение с помощью расширения scaffold для чтения занимает очень много времени. Если расширенный каркас не может быть присоединен к другому каркасу после нескольких раундов расширения, возможно, стоит протестировать дополнительный каркас (возможно, небольшой, поэтому его легко пропустить при объединении) путем поиска перекрытий в полном метагеноме.Иногда сбой расширения каркаса происходит из-за отсутствия парных чтений, которые могут быть обнаружены в конце другого фрагмента. Если они указывают, но последовательности не могут быть соединены из-за перекрытия концов, в объединенную последовательность может быть вставлен разрыв каркаса (может потребоваться обратное дополнение одного из каркасов) ». Это означает, что иногда можно связать все каркасы чернового MAG в кольцевой геном путем удлинения каркаса с последующей сборкой расширенных каркасов на основе перекрытия.Однако следует отметить, что ручное наращивание каркаса требует много времени и часто не приводит к циклической последовательности генома MAG. Дополнительные тесты должны быть выполнены для проверки точности конечного продукта, как мы описали в нашем исследовании. Как можно наращивать строительные леса? Мы вручную расширяем строительные леса в заданном контейнере, чтобы их можно было курировать с помощью Geneious. Для этого есть несколько шагов, включая (1) отображение считываний на каркасы, составляющие корзину, (2) визуализацию в Geneious и (3) ручное расширение в Geneious.Обратите внимание, что считывания, используемые для расширения, должны быть соответствующим образом размещены относительно их уже сопоставленной пары в правильной ориентации, а первоначально сопоставленная пара должна поддерживать согласованную последовательность на концах каркаса. В следующих разделах подробно описывается каждый из этих трех шагов. (1) Считывает отображение Картирование чтения может быть выполнено с помощью доступных инструментов, например Bowtie 2. Здесь мы проиллюстрируем использование первого бункера генома (содержащего один каркас), описанного в разделе «Примеры из практики, иллюстрирующие курирование черновиков MAG» в CGM, i .е., ALT_04162018_0_2um_scaffold_13. Каркас был собран из парных чтений (ALT_04162018_0_2um.1.fastq.gz и ALT_04162018_0_2um.2.fastq.gz) и имеет формат FASTA. Для чтения отображения с помощью Bowtie 2. Сначала создайте базу данных для сопоставления. Во-вторых, сопоставьте чтения с эшафотом Примечание: «-p 6» - это количество ядер, используемых для сопоставления, «-X 2000» - это наибольшая длина вставки, разрешенная при сопоставлении считываний на парном конце, которая может быть изменена на основе исследования. (2) Визуализация в генеосе Подготовьте файл каркаса (.fasta) и файл сопоставления Bowtie 2 (.sam), используя функцию в Geneious, «Файл» -> «Импорт» -> «Из файла». После импорта файла fasta выберите его и импортируйте файл SAM. Выберите соответствующую последовательность .fasta, используемую для сопоставления: Генерируются два файла, один (средний на скриншоте ниже) показывает чтения, которые сопоставлены с каркасом, другой (нижний) включает неразмещенные чтения из пар (означает, что другие чтения в этих парах были сопоставлены с эшафот через Bowtie 2). Ниже приведен обзор всех считываний, сопоставленных с каркасом, профиль покрытия обычно ровный (за исключением области между 700000 и 800000 п.о., которая является областью профага). (3) Ручное выдвижение строительных лесов Следующим шагом будет расширение каркаса вручную в Geneious. Сначала выберите файл FASTA и файл чтения без места (упомянутый выше) и используйте функцию Geneious «Выровнять / собрать» -> «Сопоставить с эталоном». Для чувствительности используйте «Custom Sensitivity» и установите «Maximum Mismatches Per Read» равным 2%, не изменяйте никакие другие параметры.Вы можете установить флажок «Сохранить список неиспользованных чтений», чтобы сохраненные чтения использовались для следующего запуска расширения. На этом шаге генерируются два файла (внизу снимка экрана ниже): один содержит 605 неразмещенных операций чтения Bowtie 2, сопоставленных с каркасом, и другой, содержащий 3284 операций чтения, которые остаются не сопоставленными. Проверьте показания, отображаемые на концах каркаса. Консенсусные последовательности чтений, которые только частично сопоставлены с каркасом, должны использоваться для расширения (см. Левый и правый концы ниже: скопируйте и вставьте последовательность чтения в строку согласованной последовательности над ними). Перед расширением каркаса считывания в парах, которые уже сопоставлены с каркасом, должны быть проверены, чтобы увидеть, размещены ли они на подходящем расстоянии, учитывая информацию о размере библиотеки. Это можно сделать, выполнив поиск по названиям чтений. Например, имя самого нижнего читаемого на правом конце - «SNL153: 233: HYKNYBCX2: 1: 1109: 2879: 14634/1 (перевернуто)», которое можно скопировать, как показано на скриншоте ниже. Таким образом, мы должны искать "SNL153: 233: HYKNYBCX2: 1: 1109: 2879: 14634/2" в исходном файле SAM от Bowtie 2, чтобы проверить его местоположение и ориентацию на карте (см. Ниже). Расширенная последовательность скаффолда может быть сохранена как «ALT_04162018_0_2um_scaffold_13_extended» (длина от 1 128 909 до 1 129 134 пар оснований). Затем сопоставьте набор неиспользованных операций чтения с последнего запуска сопоставления с расширенным шаблоном. Это следует повторять до тех пор, пока не перестанут набираться неразмещенные чтения. При необходимости полный набор данных чтения метагенома можно переназначить на расширенную последовательность каркаса, чтобы продолжить процесс. Вы можете увидеть вариации в чтениях, которые соответствуют концам каркаса, что может указывать на отдельные пути или варианты последовательности (возможно, из-за присутствия субпопуляций в окружающей среде).Как показано выше, можно определить правильный путь, если только один из двух вариантов имеет парное чтение, которое поддерживает консенсус. Если это не так, вы можете выбрать вариант пути с наиболее подходящим покрытием (или большинством чтений) или прекратить усилия по расширению. После раунда расширения каркаса может оказаться возможным найти каркасы в бункере генома, которые можно объединить (в некоторых случаях, охватываемых парными считываниями). Если каркас не найден, последовательность новых концов каркаса можно искать по всему набору метагеномных каркасов, чтобы найти фрагмент, который не был включен в корзину (с правильным содержанием GC, покрытием и филогенетическим профилем).Эту последовательность можно использовать для расширения каркаса и переназначения чтения для дальнейшей проверки. Закрытие зазора Когда скаффолдинг является этапом сборки de novo (ассемблеры, такие как IDBA_UD, metaSPAdes) данного метагеномного набора данных, N вставляются между контигами, охватываемыми чтениями с парным концом. CMAG представляют собой геномы без N, поэтому промежутки N в каркасах должны быть заполнены соответствующей последовательностью. Как можно закрыть гэп? Первый шаг должен состоять в том, чтобы проверить, действительно ли промежуток N заполнен парными чтениями (в противном случае сборка может быть химерной).Второй шаг должен заключаться в том, чтобы увидеть, присутствует ли уже последовательность в области гэпа, но дублируется ли она по обе стороны от Ns (таким образом, разрыв может быть закрыт путем удаления дублированной области и Ns). Закрытие зазора может быть выполнено автоматическими инструментами, такими как Gapfiller (Nadalin et al. 2012), даже если мы не оценивали его производительность. Здесь мы описываем альтернативную стратегию и показываем, как это можно сделать с помощью Geneious. Для иллюстрации мы используем еще один каркас из того же образца, что и описанный выше, т.е.например, ALT_04162018_0_2um_scaffold_30, который содержит зазор в лесах (эти зазоры можно быстро найти, выполнив поиск «Ns»). На рисунке ниже показаны профили сопоставления чтения из Bowtie 2. Первым шагом к устранению разрыва в строительных лесах является его устранение путем добавления подходящего числа «-» в последовательность согласования (серая линия на скриншоте ниже показывает правильное место для добавления «-»). Вот как выглядит после добавления «-» слева от области смешения. Затем мы должны переместить чтения в этой области в соответствующую сторону: Затем удалите столбцы без базы. Как вы можете видеть на скриншоте выше, есть последовательности, общие для операций чтения, которые можно скопировать и вставить в последовательность шаблонов, чтобы частично заполнить пробел. Несмотря на то, что считывания в областях гэпов обладают высоким сходством последовательностей, мы также можем видеть некоторые варианты с одним нуклеотидом (SNV). Мы выбираем согласованные последовательности, разделяемые большинством прочтений. Неподдерживаемая консенсусная последовательность должна быть заменена на Ns. Затем мы сохраняем измененный каркас как «ALT_04162018_0_2um_scaffold_30_modified». Мы сопоставляем неразмещенные парные чтения (такие же, которые используются для удлинения концов каркаса, см. Выше) с "средней чувствительностью / быстротой" на модифицированный каркас. Выходной профиль отображения области с зазором строительных лесов показан ниже: Откройте разрыв и отсортируйте показания, как описано выше. Добавьте согласованные последовательности, разделяемые чтениями, в каркас.Не беспокойтесь о SNV; мы разберемся, что происходит, когда разрыв будет ликвидирован. Выполнение сопоставления с неразмещенными парными считываниями до устранения разрыва: Профили сопоставления указывают на то, что разрыв будет закрыт после удаления оснований в каркасах без поддержки чтения: Затем сопоставьте чтения с каркасом с закрытым зазором, не допуская несоответствия (настраиваемая чувствительность, допустимое нулевое несоответствие), чтобы увидеть, покрыта ли область повсюду. Закрывая пробел, мы получаем полную аминокислотную последовательность для соответствующего гена, что подтверждается поиском BLASTp: Выводы В этом блоге мы покажем, как расширение строительных лесов и закрытие промежутков между ними может быть выполнено в Geneious.Мы надеемся, что эта информация поможет пользователям улучшить качество своих MAG. Важно отметить, что в будущем необходимо разработать автоматические инструменты для выполнения этих этапов курирования. Пожалуйста, не стесняйтесь оставлять комментарии ниже или связываться с нами по электронной почте, если вы работаете над автоматизированными решениями или у вас есть разные стратегии или советы по улучшению MAG. Подход каркаса, который мы предлагаем, основан на наличии общих гаплотипов в геномах вида.Таким образом, в принципе, он использует эффект неравновесия по сцеплению (LD) (Lewontin and Kojima 1960). Каждое отдельное растение, которое сравнивается с эталонным геномом, может показывать либо эталонный аллель, либо не эталонный аллель в данном геномном положении. Поскольку вариации наследуются внутри вида, мы ожидаем, что группы особей покажут нереференсные аллели в одних и тех же референсных положениях генома. В зависимости от размера анализируемой области генома мы находим ряд таких групп, каждая из которых представляет один гаплотип в этой области (рис.1). В популяции картирования, где родительские гаплотипы различаются известным набором вариантов положений (или любым набором маркеров), можно отнести каждую область геномов следующего поколения к одному из родительских гаплотипов. В этом исследовании, глядя на людей, которые не связаны напрямую, мы не знаем, сколько «начальных» гаплотипов следует учитывать, чтобы определять гаплотипы просто на основе общих вариаций в данной области генома. Другими словами, ожидается, что в любой небольшой области генома будут представлены две группы индивидуумов, одна из которых будет похожа на ссылку, а другая - на другую.В более крупной области генома более двух групп могут стать очевидными по мере рассмотрения большего числа позиций. Такое группирование индивидуумов на основе общих вариантов приводит к паттерну вариаций для каждой позиции генома в геноме. Ожидается, что переход от одного паттерна к другому произойдет через более длинные геномные расстояния (соответствующие ожидаемым расстояниям для мейотического кроссинговера), так что последовательности фрагментированной сборки генома, используемые в качестве эталона, могут демонстрировать паттерны вариаций, которые продолжаются через пробелы в сборке.Таким образом, паттерны непрерывных вариаций (то есть варианты позиций, которые группируют индивидов подобным образом) могут служить руководством для упорядочивания и ориентации собранных контигов. Позиции генома, которые не показывают каких-либо вариантов или зашумленных позиций, должны быть удалены, а паттерны вариаций необходимо суммировать, чтобы уменьшить количество паттернов, которые нужно сравнивать между контигами. Варианты 22 человек (1-22) по сравнению с эталонной сборкой (Ref), показаны только варианты положения (схематично).В геномном интервале A можно выделить два гаплотипа, представленных индивидами 1–12 и 13–22 соответственно. В геномном интервале B можно выделить три гаплотипа, представленных индивидами 1–5, 6–12 и 13–22 соответственно. Во всем геномном регионе (C) можно выделить четыре гаплотипа, представленных индивидами 1–5, 6–12, 13–16 и 17–22 соответственно. Всего существует три различных паттерна вариации для каждой позиции в геноме: первый паттерн в интервале A, второй в интервале D, третий в интервале E Основными этапами являются: секвенирование ряда особей, считывание карт по эталону. сборка, вариант, требующий для каждого индивидуума по сравнению с эталоном, фильтрация неинформативных или зашумленных геномных позиций, обобщение паттернов вариаций в пределах геномных интервалов и сравнение обобщенных паттернов вариаций между контигами. Для обработки сопоставимых геномных единиц собранные контиги делятся на сегменты фиксированного размера, а модели вариаций суммируются и подсчитываются для каждого сегмента (рис. 2). Редкие паттерны (т.е. встречающиеся только в одной или двух геномных позициях внутри сегмента) удаляются, а оставшиеся обобщенные паттерны вариаций упоминаются как «отпечатки пальцев» для каждого сегмента. Сравнение контигов основано на сравнении таких вариаций отпечатков пальцев, и контиги упорядочиваются в соответствии с их общими отпечатками.Контиги объединяются и / или организуются в каркасы, если доступны вещественные доказательства, например, предоставленные поддерживающими парами матов или длинными чтениями. Полагаться исключительно на вещественные доказательства, такие как пары полов, часто неоднозначно из-за больших повторяющихся областей. Эти области либо не будут охватываться чтениями пар сопряжения, либо будут охвачены чтениями, имеющими несколько позиций выравнивания. Следовательно, информация, основанная на гаплотипах, обеспечивает дополнительное руководство и подтверждение для скаффолдинга. Два геномных сегмента сгруппированы и сжаты для получения их вариаций отпечатков пальцев.Варианты (красный: гомозиготный, фиолетовый: гетерозиготный), референсные аллели (синий) и отсутствующая информация (серый) указаны для каждого из 50 образцов квиноа (столбцы) вдоль геномной области 20 т.п.н. в сборке Qpac (tig00006496, 1,75 Мбит / с. до 1,77 Мбит / с). Показаны только позиции, несущие альтернативные аллели (строки). Все рассматриваемые позиции ( a ) фильтруются ( b ) для сохранения информативных позиций, где по крайней мере половина образцов покрывает позицию по крайней мере с одним считыванием и не более 300 считываний, и где есть по крайней мере две ссылки. и два альтернативных генотипа.Вставки и удаления отбрасываются, и сохраняются только двуаллельные SNP. Последовательность генотипов в позиции называется паттерном вариации. Паттерны группируются посредством попарных сравнений ( c ) и сжимаются ( d ) для формирования вариационного отпечатка пальца. Подобные паттерны вариаций сохраняются, если они встречаются по крайней мере в трех геномных позициях в пределах сегмента. Чтобы их считали похожим паттерном, они не должны иметь различий в гомозиготных вариантах и могут иметь до 10% различий в своих гетерозиготных вариантах.Отпечатки пальцев всех сегментов в геноме сравниваются друг с другом, чтобы найти общие модели вариаций между разными сегментами ( e ) В качестве доказательства концепции мы воспользовались ранее генерировали данные генотипирования на основе полногеномного секвенирования 1135 образцов модельного растения Arabidopsis thaliana (Alonso-Blanco et al., 2016). В контексте этого проекта была определена однонуклеотидная вариация относительно эталонной последовательности A.thaliana Колумбийский (Col-0) генотип. На основе этой информации о вариациях мы определили паттерны вариаций для каждой позиции в эталонной сборке Col-0 и сгенерировали отпечатки вариаций на сегмент размером 100 кбит / с. Количество общих отпечатков пальцев между такими сегментами 100 кбит / с сравнивалось для наборов от 15 до 100 случайно выбранных образцов из 1135 доступных наборов данных (Таблица 1). Всего с 15 случайно выбранными образцами мы обнаружили в среднем по геному три общих отпечатка пальца на мегабазу, подтверждающие сборку (т.е.е., расположенных в соседних сегментах) и до 0,30 общих отпечатков пальцев на мегабазу между удаленными сегментами, не поддерживающими структуру сборки (таблица 1, онлайн-ресурс 1). Большинство сегментов, которые были правильно идентифицированы как смежные на основании общих отпечатков пальцев, были расположены в центромерных регионах. При использовании 100 случайно выбранных образцов количество общих отпечатков пальцев в соседних сегментах увеличилось до 16 на мегабазу, распределенных по всему геному, с не более чем 1,90 удаленных родственников на мегабазу.Использование наборов данных генотипирования из 50 случайно выбранных образцов привело к получению 11-13 смежных общих отпечатков пальцев и 1,4-2 удаленных общих отпечатков пальцев на мегабазу. Таким образом, сбор информации о гаплотипах на основе 50 повторно секвенированных образцов оказался хорошим компромиссом между количеством секвенированных особей, количеством отпечатков пальцев, правильно связывающих соседние сегменты друг с другом, и частотой ложноположительных результатов удаленных связей (рис. 3). Гаплотипические связи в геноме A. thaliana , полученные с различными наборами данных. Каждая точка представляет собой геномный сегмент размером 100 т.п.н. Синие ссылки: внутрихромосомные отношения; красные ссылки: межхромосомные или внутрихромосомные отношения с расстоянием> 1 Мбит / с. Координаты хромосом отображаются в Мбит / с с интервалом 5 Мбит / с. Отношения основаны на наборах данных SNP с частотой минорных аллелей ≥ 0,05. Каждая дуга соответствует А.thaliana , где отношения показаны на основе информации о гаплотипах с 15 ( a ), 30 ( b ), 50 ( c ) и 100 ( d ) образцами, выбранными случайным образом. Последняя выборка ( e )
основан на 50 образцах северной Швеции, а также имеет самую высокую плотность связей на основе гаплотипов Затем мы оценили влияние генетического родства образцов на обнаружение смежных геномных сегментов на основе общих отпечатков пальцев.Для этого мы выбрали образцы из двух различных географических областей вместо случайного отбора. Пятьдесят итальянских образцов показали плотность смежных общих отпечатков пальцев (11,5 на мегабазу), аналогичную случайно выбранным. Однако при выборе 50 образцов из Швеции мы наблюдали более чем в два раза больше (21,9) смежных общих отпечатков пальцев на мегабазу (таблица 1). Набор шведских образцов включал образцы со всей страны (с севера и юга), при этом северные образцы показали самые низкие локальные попарные генетические дистанции из любых A.thaliana было отобрано образца в Европе, и у него также был самый высокий уровень редких аллелей в контексте всего набора данных по 1135 образцам (Alonso-Blanco et al., 2016). Южно-шведские образцы имели более высокие попарные расстояния по сравнению с северными, но также имели меньше редких аллелей. Таким образом, образцы из северной Швеции имели наибольшее количество общих гаплотипов. Это также был набор данных с лучшей производительностью с точки зрения плотности отношений на основе гаплотипов. Образцы из Италии были выбраны на основе географической близости в пределах северной Италии, но известно, что они сгруппированы с различными популяциями, распределенными по всей Евразии (Alonso-Blanco et al.2016). Тесное генетическое родство среди шведских образцов было отражено в количестве вариантов: было меньше различий между шведскими образцами A. thaliana по сравнению с эталоном Col-0 по сравнению с случайно выбранными образцами (Таблица 1). Например, в 50 шведских образцах было 2,2–3,2 миллиона позиций вариантов по сравнению с 3,9–4,4 в 50 случайно выбранных образцах. Это подчеркивает важность наличия консервативных гаплотипов на больших геномных расстояниях, а не просто большого количества вариантов для увеличения количества смежных общих отпечатков пальцев. Мы использовали информацию о гаплотипах для создания основы сборки de novo генома квиноа, который примерно в десять раз больше, чем геном A. thaliana. В больших геномах, богатых повторами, упорядочение контигов и создание каркасов становится сложной задачей. В качестве эталонного генома мы секвенировали геном квиноа боливийского образца CHEN125, типичного белосемянного сорта, первоначально собранного в Лауачака в боливийском регионе Альтиплано на высоте 3800 м.Секвенирование было выполнено на платформе секвенирования Pacific Biosciences RS II, в результате был получен набор данных со средней длиной считывания 10464 п.н. и 65-кратным геномным покрытием (таблица 2), предполагая, что размер гаплоидного генома составляет 1,45 Гбит / с (Kolano et al. 2012). Распределение длины чтения имело два пика: 6800 пар оснований и 16000 пар оснований (Интернет-ресурс 2). Необработанные данные были скорректированы и собраны с использованием конвейера Canu (Koren et al., 2017), в результате чего размер сборки составил 1,32 Гбит / с, состоящий из 6747 контигов с длиной N50 608 кбит / с (называемой сборкой «Qpac»).Пятьдесят процентов сборки Qpac находилось в 546 контигах (таблица 3). Из набора из 2121 высококонсервативных генов растений, используемых в качестве входных данных для BUSCO (Seppey et al.2019; Simão et al.2015), мы обнаружили 2050 генов (96,7%) в Qpac, что предполагает полный охват пространства генов в нашей сборке (Таблица 4). Из 2050 BUSCO 184 были обнаружены более чем в двух экземплярах (до шести копий), а 1287 BUSCO появились ровно дважды. Было дуплицировано около 70% генов BUSCO, что отражает тетраплоидную природу генома квиноа.Действительно, несмотря на сборку диплоидного генома, квиноа произошла в результате гибридизации двух видов Chenopodium . Гомеологичные последовательности были успешно собраны по отдельности, что отражено в размере сборки 1,32 Gbp, что соответствует двойному размеру гаплоида его родительских доноров генома. Следовательно, большинство групп BUSCO было обнаружено дважды, поскольку они присутствуют в отдельных гомеологичных хромосомах. Мы сопоставили данные парных концов Illumina от того же образца сборки PacBio и обнаружили, что 450 353 сайта были гетерозиготными (включая индели и SNP), что соответствовало 0,034% от общего количества. геном. Мы оценили размер генома квиноа образца CHEN125 на основе данных секвенирования с учетом частотного распределения k-мер. Основываясь на 17-мерах, мы оценили размер диплоидного генома в 1.22 Gbp (Интернет-ресурс 11). Аналогичные результаты были получены для оценок на основе значений от k = 19 до k = 27 (от 1,23 до 1,27 Гбит / с). Квиноа - аллотетраплоидное растение, но его субгеномы достаточно различаются, так что его геном по существу ведет себя как диплоид (Schiavinato et al. 2021). Расчетный размер генома 1,22 Гбит / с и размер сборки 1,32 Гбит / с хорошо согласуются. Для того, чтобы связать собранные контиги с 18 хромосомами квиноа, мы загрузили последовательности, которые ранее использовались в качестве маркеров для создания генетической карты генома квиноа на основе однонуклеотидной вариации ( Maughan et al.2012) и расположил их в сборке Qpac. Из 511 доступных маркеров, определяющих 29 групп сцепления, 356 выровнены однозначно с контигами Qpac. Всего 282 контига Qpac, составляющих 270 Мбит / с (20,5% сборки Qpac), можно однозначно отнести к группе сцепления. Из этих контигов 225 контигов содержали один маркер, а 57 контигов содержали от двух до пяти маркеров (Интернет-ресурс 3). Доля хромосомно присвоенных контигов может быть существенно увеличена после повторного секвенирования набора из 50 общедоступных образцов квиноа для определения вариантов и обнаружения гаплотипов: когда гаплотипы охватывают несколько контигов, могут быть установлены связи с еще не присвоенными контигами (см. Ниже). Мы выбрали 50 образцов квиноа из Боливии и Перу, происходящих из окрестностей озера Титикака, для полногеномного секвенирования с низким охватом (Интернет-ресурс 4). Посевной материал этих образцов доступен в общедоступных хранилищах, так что образцы могут быть повторно выращены при необходимости. Секвенирование геномной ДНК было выполнено на платформе Illumina, что дало в среднем 32 миллиона пар считывания на образец, что в среднем составляет 5.7-кратный геномный охват (от 4,3 до 7,4) до фильтрации. Качественные отфильтрованные чтения (средний охват от четырех до семи раз) от образцов были сопоставлены с эталонным геномом при средней скорости картирования от 47 до 59% (однозначное сопоставление пар в пределах ожидаемого размера вставки с геномом). Первоначальный вызов варианта перед дальнейшей фильтрацией с использованием Qpac в качестве эталона привел к получению в общей сложности 5 005 967 позиций вариантов, из которых 596 685 были вставками или делециями (инделциями) и 4 409 282 были однонуклеотидными полиморфизмами (SNP) (Интернет-ресурс 5 ) .Это было 1,44 миллиона вариантов на образец, в среднем 62% из которых были гомозиготными, а 38% - гетерозиготными. Для сравнения мы применили тот же подход к общедоступной сборке квиноа чилийского сорта QQ74 (Jarvis et al., 2017), называемого ASM168347v1, что дало 594194 инделя, 4706603 SNP и 1,89 миллиона позиций вариантов в среднем на образец (68% гомозиготный, 32% гетерозиготный). Учитывая, что сборка ASM168347v1 основана на сорте квиноа из Чили, в то время как эталон Qpac и повторно секвенированные образцы происходили как из боливийского, так и из перуанского региона Альтиплано, ожидалась более высокая степень вариации повторно секвенированных линий по сравнению с ASM168347v1. .Немного меньшее количество инделей в ASM168347v1 можно объяснить более низкой скоростью сопоставления с этим геномом (на 0,5–1% меньше) и большим количеством считываний, отображаемых как единичные считывания (т. Е. На 1–2,5% больше считываний, которые не могли быть выровнены как пары из-за одно из сопряжений не соответствует ожидаемому диапазону размеров вставки). Это может указывать на структурные изменения, которые трудно обнаружить с помощью коротких данных секвенирования парных концов Illumina. Коэффициент перехода / трансверсии вариантов, обнаруженных с использованием любого эталона, имел одинаковое значение 1.50. Пять образцов с наибольшим числом вариантов были одинаковыми для обоих геномных ссылок и происходили из провинции Акомайо (Перу). Акомайо расположен в регионе Куско, который граничит с регионом Аякучо, где предполагается, что одомашнивание квиноа произошло 5000 лет назад (Lumbreras et al. 2008). Средняя плотность вариации от 3,34 положений на т.п.н. (CHEN125) до 3,53 положений на т.п.н. (QQ74) в геноме квиноа сравнима с плотностями вариантов у других одомашненных растений (Xu et al.2019) и намного ниже, чем у дикорастущих растений, таких как A. thaliana (101,95 позиций на тыс. П.о.) (Alonso-Blanco et al., 2016). Было 3589483 позиции вариантов в Qpac, если рассматривать только двуаллельные SNP, покрытые по крайней мере половиной из 50 человек и с частотой минорных аллелей> 5%, что было набором данных SNP, используемых для дальнейшего анализа в контексте гаплотипов. строительные леса. С этими настройками мы наблюдали 1,18 миллиона позиций вариантов в среднем на образец, из которых 61% были гомозиготными и 39% были гетерозиготными. Варианты по собранным контигам Qpac анализировали в сегментах размером 100 кб. Каждый контиг меньше 100 кбит / с (4390 контигов на общую сумму 185 Мбит / с) считался одним сегментом. Всего для нашей сборки 1,32 Гбит / с мы получили 15 204 таких сегмента. Для каждого сегмента мы определили вариант отпечатка пальца и провели полное сравнение отпечатков пальцев для каждого сегмента. Некоторые сегменты не отображали вариаций и, следовательно, не имели отпечатков пальцев.Первоначально было 12366 сегментов, распределенных по 3172 контигам, показывающих общие паттерны вариаций, по крайней мере, с одним другим сегментом. Были удалены очень распространенные паттерны, а также паттерны, которые поддерживались только несколькими позициями внутри сегмента (предположительно возникающие из-за редких мутаций, недавних мутаций или неправильных вызовов вариантов). Отпечатки пальцев между сегментами должны были разделять по крайней мере 10% их вариантных паттернов, чтобы их можно было рассматривать как достаточно поддерживаемые отношения на основе гаплотипов. После фильтрации слабо поддерживаемых отношений сегментов было найдено 2188 контигов, которые имели общие отпечатки вариаций, по крайней мере, с одним другим контигом.Такие контиги образовали 371 группу, состоящую от двух до 82 контигов (Интернет-ресурс 6). Всего 2188 контигов составили 907 Мбит / с, то есть 69% сборки. Поскольку отпечатки вариаций могут охватывать несколько контигов, локальный порядок контигов не всегда можно было разрешить. Среди 371 группы 306 сформировали небольшие соединительные сети, которые можно было однозначно разделить на линейную структуру (рис. 4). Эти 306 групп содержали 849 контигов (2–11 контигов на группу) и охватывали 350 Мбит / с. Сети оставшихся 65 групп, состоящих из 1339 контигов, были более сложными (Интернет-ресурс 7).Чтобы получить окончательную сборку RefCHEN125, мы только скаффолдили или объединили контиги, для которых были доступны фактические вещественные доказательства, т. Контиги, которые не могли быть физически связаны, оставались в группе контигов (что отражено в названии последовательностей сборки). В этих случаях информация о гаплотипах не позволяет определить точный порядок контигов, а просто предоставляет информацию об их смежности.По нашему определению, группы контигов могут состоять из истинных каркасов , но могут также содержать контиги без точной информации о местоположении. Порядок и ориентация четырех контигов Qpac на основе информации о гаплотипах и вещественных доказательств. Линии между контигами указывают на отношения высокой достоверности, основанные на общих моделях вариаций. Картирование мат-пар (соединительные линии справа, указано количество мат-пар) подтвердило смежность контигов и выявило их окончательный порядок и ориентацию; tig00010021 пришлось перевернуть в соответствии с совпадающим расположением пар матов.Результирующий размер каркаса составлял 561 kbp последовательности плюс 28 kbp пробелов Среди групп, связанных вариационными отпечатками пальцев, 123 содержали контиги, ранее привязанные к генетическим маркерам (Интернет-ресурс 6, Интернет-ресурс 3). Было достигнуто общее согласие между генетическими маркерами и группами, сформированными по вариационным отпечаткам пальцев. Например, пять из 38 контигов, расширяющих контиг «tig00000000» 3,1 Мбит / с, ранее привязанный к группе сцепления 2 (LG 2), несли генетические маркеры, все из которых последовательно были из LG 2.Из всего 282 генетически заякоренных контигов Qpac (270 Мбит / с) было 214 контигов, которые можно было расширить с помощью 1206 дополнительных контигов (452 Мбит / с) с помощью общих отпечатков вариаций (таблица 5). Остальные группы, не привязанные к генетическим маркерам, охватывают 250 Мбит / с в 768 контигах в 248 группах. Таким образом, из 1.Сборка Qpac из киноа 32 Гбит / с, 908 Мбит / с продемонстрировала улучшенное упорядочение после использования общих отпечатков вариаций (т. Е. Информации о гаплотипе). В оставшихся 4559 контигах (411 Мбит / с) со средним размером 90 Кбит / с отсутствовали вариации, поэтому сравнение отпечатков пальцев не могло быть выполнено. Однако из них 68 контигов в 65 Мбит / с уже были заякорены в хромосоме на основе генетических маркеров. Общая генетически заякоренная фракция сборки после рассмотрения информации о гаплотипе составила 722 Мбит / с, что составляет 55% сборки Qpac. Порядок контигов, основанный на информации о гаплотипах, был проверен на основе согласованности с парами партнеров Illumina, длинных чтений и / или перекрывающихся концов контигов, по сравнению с другой сборкой квиноа и назначением для связывания групп с помощью генетических маркеров. Среди 123 групп контигов, которые также содержали по крайней мере один генетический маркер, только три группы содержали маркеры из двух разных хромосом. Так было с контигом Qpac «tig00007425» (965 кб), ранее назначенным LG 1, но связанным с несколькими контигами, назначенными LG 6 по информации гаплотипа.Однако «tig00007425» также содержал уникальный выравнивающий генетический маркер из LG 6 с более низкой оценкой картирования (Интернет-ресурс 3). Маркер LG 6 Cq04033_520 сопоставлен близко к левому концу контига, а маркер LG 1 Cq07822_1014 сопоставлен рядом с правым концом контига. Все контиги, связанные с помощью вариационных отпечатков пальцев и присвоенные LG 6, выходят из левого конца «tig00007425». При осмотре покрытия для чтения мы заметили падение покрытия между двумя маркерами, указывающее на возможное место неправильной сборки в «tig00007425.«Подобные случаи были обнаружены в двух других группах, содержащих противоречивые назначения групп сцепления; эти группы также показали падение покрытия между противоречивыми маркерами («tig00005716», присвоенный LG 12, сгруппированный с четырьмя назначенными контигами LG 15, и «tig00008618», назначенным LG 3, сгруппированным с двумя назначенными контигами LG 10). Таким образом, помимо подтверждения доступных присвоений LG, информация о гаплотипах привела к идентификации трех случаев потенциальных неправильных сборок, которые были разрешены путем разделения соответствующих контигов.С другой стороны, 36 групп контигов на основе гаплотипов содержали по крайней мере два генетических маркера из одной и той же LG и подтверждали друг друга. Группа, содержащая наибольшее количество контигов, включала два контига LG 1 и всего 82 контига (всего 13 Мбит / с, онлайн-ресурс 6). Самая большая группа с точки зрения базовых пар включала 13 контигов на LG 5 и включала в общей сложности 74 контига (40 Мбит / с). В этой группе также было наибольшее количество контигов, содержащих генетические маркеры, то есть всего 13 маркеров.В целом организация контигов на основе информации о гаплотипах подтвердила назначение LG на основе генетических маркеров. Группы контигов в целом соответствовали сборке квиноа на хромосомном уровне ASM168347v1 чилийского сорта QQ74 (Jarvis et al. 2017). Например, при выравнивании всех контигов и групп контигов, относящихся к LG 2 (всего 44 Mbp), со сборкой ASM168347v1, большинство из них имело выравнивание последовательностей от конца к концу в хромосоме 2 (рис. 5). Хромосома 2 ASM168347v1 имеет длину 59 Мбит / с, включая 2 Мбит / с неуказанных оснований.В то время как сети, созданные с помощью информации о гаплотипах, не могли полностью определить порядок и ориентацию каждого контига, общий порядок более крупных контигов и их хромосомная принадлежность были последовательно подтверждены ASM168347v1. Каркас CHEN125 позволил выделить три крупномасштабные инверсии в хромосоме 2 по сравнению с QQ74, что было сильно подтверждено данными о парных концах (рис. 5, синие линии). Выравнивание нукмеров между последовательностями RefCHEN125, присвоенными хромосоме 2 квиноа, и сборке хромосомы 2 ASM168347v1.Синие линии изображают крупномасштабные инверсии, а красные линии изображают большие участки выравнивания по всей длине между двумя сборками Для дальнейшей проверки порядка и ориентации контигов на основе гаплотипов мы упорядочили пары партнеров Illumina с пятью различными размерами пролета в диапазоне от 2,5 кбп до 8,1 кбп (Интернет-ресурс 8). Мы также приняли во внимание перекрытия между контигами и охватом длинных чтений. Среди всех отношений, основанных на гаплотипах (6435), 3351 имел по крайней мере одно поддерживающее долгое чтение. Длинные чтения имели размер, аналогичный размеру самой большой вставки библиотеки пар сопряженных пар (около 10 кбайт), так что соединения между контигами, перекрывающимися более чем на 10 кбайт, или между контигами, у которых неправильно собранные концы длиннее 10 кбайт, не могли быть охвачены чтениями.Всего мы соединили 1848 контигов в 790 последовательностей, содержащих от 2 до 13 контигов. Всего 1298 соединений между контигами были подтверждены парами спаривания, 416 соединений были подтверждены перекрытиями и 712 соединений были подтверждены как парами спаривания, так и перекрытиями. Кроме того, длинные чтения подтвердили 1480 соединений, уже поддерживаемых парами сопряжения или перекрытиями. В целом, из 2188 контигов, связанных с помощью информации о гаплотипах, 84,5% имели подтверждающие физические доказательства в виде мостовых пар, совпадающих длинных считываний и / или перекрытий для установления их связей. Мы интегрировали информацию из перекрытий, пар сопряжений и длинных считываний, чтобы улучшить непрерывность сборки Qpac, и получили версию каркаса под названием RefCHEN125. Сборка RefCHEN125 имеет длину N50 1079 т.п.н., общий размер 1,314 Гбит / с и включает 5689 последовательностей (таблица 3). Разница в размере между RefCHEN125 в отношении Qpac была в основном из-за перекрытия контигов, поддерживаемого парами сопряжения или длинными чтениями. Из-за размеров диапазона наших библиотек сопряженных пар (макс.размер пика 8 кб), которые находились в том же диапазоне, что и длина субпотока PacBio, обнаруженные ссылки чаще приводили к слиянию, чем к вставке пропусков. Основное улучшение произошло за счет переупорядочения 2188 контигов в соответствии с их смежностью в геноме, выведенной из информации о гаплотипах. Сборка Qpac первоначально предоставила непрерывную информацию для максимального участка в 7,3 Мбит / с, тогда как RefCHEN125 предоставил основанные на гаплотипах доказательства упорядоченных контигов в больших регионах до 39,7 Мбит / с и скаффолдинговых контигов в регионах до 10.5 Мбит / с. Последовательности в файле FASTA RefCHEN125 были упорядочены и переименованы, чтобы отразить их положение в собранном геноме. //www.ensembl.org/?mobileredirect=no Геномные сборки являются иерархическими.Самыми короткими компонентами сборки являются контиги, которые представляют собой последовательности, взятые у отдельных лиц. Контиги собираются в более длинные каркасы, а каркасы собираются в хромосомы, если имеется достаточная информация о картировании. Многие сборки генома собраны только до уровня каркаса. Строительные леса подразделяются на три категории: Отношения между контигами, каркасами и хромосомами определены в файлах AGP. Эти файлы описывают, как собранные последовательности (например, хромосомы) составляются из их компонентов (например, каркасов). В Ensembl мы импортируем последовательность ДНК на уровне контигов в наши основные базы данных. Мы также импортируем файлы AGP для сопоставлений контиг-скаффолд, контиг-хромосома и скаффолд-хромосома.Это позволяет нам генерировать последовательность каркаса и хромосомы на лету, сшивая последовательности контигов вместе, как указано в файлах AGP. Для каждой сборки генома мы определяем набор последовательностей верхнего уровня. Это участки последовательности в сборке генома, которые не являются компонентом другой области последовательности. Например, когда геном собран в хромосомы, последовательностями верхнего уровня будут хромосомы и любые нелокализованные или неразмещенные каркасы.
Рисунок 6.Удалить слабые кромки процедура remove_weak_edges (график g):
для ребер e = (u1, v2), e1 = (u1, v1), e2 = (u2, v2) в g
если g [e] .n Удалить неоднозначные кромки
Рис. 7. Удаление неоднозначных краев процедура remove_ambiguous_edges (график g):
для ребра e = (u, v) в g
если g [u] .outdeg> 1 или g [v] .indeg> 1
remove_edge (e, g)
Сборка смежных путей
bwa
bwasw -b9 -q16 -r1 -w500 . Выравнивания длиной не менее 200 п.н.
используется для расчета выровненного контига N50, исключая вторичный
выравнивания запроса, которые перекрывают большее выравнивание более чем на
50% меньшего выравнивания. Раскладки длиной не менее 500 п.н.
с качеством отображения не менее 10 использовались для идентификации
точки останова в выравнивании контигов по ссылке. ABySS ALLPATHS-LG Ферми SGA SOAP Деново Contig bp 2.70 G 2,61 г 2,75 г 2,76 G 2,66 г Выровненный контиг bp 2,69 G 2,61 г 2,73 G 2,74 G 2,64 г Крытый исх. bp 2,66 G 2,59 г 2,70 г 2,70 г 2,64 г Contig N50 9,75 к 23,8 к 16.6 к 9.91 к 11,1 к Выровненный контиг N50 9,72 к 23,7 к 16,6 к 9,90 к 11,1 к Точки останова Contig 1590 3380 920 1549 931 Подмости N50 273 к 11,5 M 446 к 167 к 565 к Не менее 500 б.п. и Q10 276 к 11.3 м 462 к 173 к 565 к Выровненные леса N50 270 к 2.30 M 458 к 168 к 557 к Точки разрыва строительных лесов 1933 3008 1333 2101 717 Программное обеспечение
Данные секвенирования образца человека NA12878
Библиотека переулок (индекс ) Читает Базы (п.н.) Solexa – 18483 20FUKAAXX100202_1 159875468 16,1 г Solexa – 18483 20FUKAAXX100202_3 1546 15,6 г Solexa – 18483 20FUKAAXX100202_5 156932536 15.9 G Solexa – 18483 20FUKAAXX100202_7 155034158 15,9 г Solexa – 18484 20FUKAAXX100202_2 149658350 15,1 г Solexa – 18484 20FUKAAXX100202_4 145881464 14,7 г Solexa – 18484 20FUKAAXX100202_6 147970372 14,9 г Solexa – 18484 20FUKAAXX100202_8 150971682 15.2 G Всего 1221224906 123,3 г Запуск SRA переулок (индекс ) Читает Базы (п.н.) SRR067773 2025JABXX100603_5 186100680 18.8 G SRR067778 2025JABXX100603_7 194515894 19,6 г SRR067779 2025JABXX100603_6 188012974 19,0 г SRR067786 2025JABXX100603_8 189208654 19,1 г Итого 757838202 76,5 г Сборки человеческого образца NA12878
Сборка строительных лесов с арками и ссылками - документация faircloth-lab e4a8853
/ home / brant / project / shared / bin / ). wget -O longranger-2.2.2.tar.gz "<длинная ссылка с веб-сайта>"
tar -xzvf longranger-2.2.2.tar.gz
mkdir 10x-diglossa-scaffold && cd $ _
mkdir longranger-ouput && cd $ _
qsub для запуска longranger и обработки чтения. Это должно занять <24 часов для заархивированных данных последовательности размером ~ 40 ГБ. Обработка в основном обрезает чтение, чтобы удалить информацию о штрих-коде и адаптере и помещает информацию о штрих-коде в заголовок fastq: #! / Bin / bash
#PBS -q checkpt
#PBS -A <распределение>
#PBS -l walltime = 24: 00: 00
#PBS -l узлов = 1: ppn = 20
#PBS -V
#PBS -N longranger_basic
#PBS -o longranger_basic.из
#PBS -e longranger_basic.err
#PBS -m abe
#PBS -M [email protected]
экспорт PATH = / home / brant / project / shared / bin / longranger-2.2.2 /: $ PATH
cd $ PBS_O_WORKDIR
longranger basic \
--id = <мое_имя> \
--fastqs = / путь / к / my / demuxed / fastq / files \
--localcores 20 1> longranger-basic.stdout 2> longranger-basic.stderr
10x-diglossa-scaffold создайте новый каталог для этих данных и создайте файл reads.fof : mkdir barcode-multiplicities && cd $ _
echo `readlink -f ../ longranger / 10x-diglossa / out / barcoded.fastq.gz`> reads.fof
reads.fof будет содержать одну строку, которая выглядит так: / ddnB / work / brant / 10x-diglossa-scaffold / longranger-output / 10x-diglossa / out / barcoded.fastq.gz
.
├── кратность штрих-кода
│ └── читает. Fof
└── longranger-output
├── 10x-диглосса
├── arks-make.orig
├── arks-make.txt
├── longranger_basic.err
├── longranger_basic.out
├── longranger.qsub
└── raw-fastq
#! / Bin / bash
#PBS -q single
#PBS -A <распределение>
#PBS -l walltime = 24: 00: 00
#PBS -l узлов = 1: ppn = 1
#PBS -V
#PBS -N arks_multiplicities
#PBS -o arks_multiplicities.из
#PBS -e arks_multiplicities.err
загрузка модуля perl / 5.24.0 / INTEL-18.0.0
экспорт PATH = / home / brant / project / shared / bin /: $ PATH
cd $ PBS_O_WORKDIR
calcBarcodeMultiplicities.pl reads.fof> read_multiplicities.csv
мкдир на подмости
# rsync до этого каталога оттуда, где расположены контиги
rsync -avLP diglossa.contigs.fasta [email protected]:/home/brant/work/10x-diglossa-scaffold/to-scaffold/
.
├── кратность штрих-кода
│ ├── multiplicities.qsub
│ ├── read_multiplicities.csv
│ └── читает. Fof
├── longranger-output
│ ├── 10x-диглосса
│ ├── arks-make.orig
│ ├── arks-make.txt
│ ├── longranger_basic.err
│ ├── longranger_basic.out
│ ├── longranger.qsub
│ └── raw-fastq
└── к эшафоту
└── диглосса.contigs.fasta
mkdir arks-scaffolded && cd $ _
make : wget -O arks-make.txt https://gist.githubusercontent.com/brantfaircloth/a714928d2824a83684254587255f4c57/raw/de6050cf759b81e87342fc521a4cfb416401b333/arks-make.
сделать -f arks-make.txt
bwa и для arks , которые называются -t (около строки 24) и - нити (около 37 строки).Вы хотите настроить эти значения в соответствии с количеством ядер в любой системе HPC, в которой вы работаете. arks не использует MPI, поэтому вы будете отправлять данные только одному узлу (таким образом, вам нужно знать, сколько процессов может запускать этот единственный узел). arks-scaffolded создайте символические ссылки на наши данные fastq и нашу сборку: ln -s ../longranger-output/10x-diglossa/outs/barcoded.fastq.gz
ln -s ../to-scaffold/diglossa.contigs.fasta
, мы можем настроить файл qsub для запуска arks, предполагая, что мы работаем на @supermike. Важно то, как arks обрабатывает ожидаемые имена файлов - убедитесь, что структурировали их правильно, иначе вы получите эту ошибку. По сути, это означает, что вам нужно обращаться к контигам без включения расширения .fasta и чтения без включения .fastq.gz расширение: #! / Bin / bash
#PBS -q checkpt
#PBS -A <распределение>
#PBS -l walltime = 72: 00: 00
#PBS -l узлов = 1: ppn = 16
#PBS -V
#PBS -N arks_scaffolding
#PBS -o arks_scaffolding.out
#PBS -e arks_scaffolding.err
# загрузить несколько модулей
загрузка модуля gcc / 6.4.0
загрузка модуля perl / 5.24.0 / INTEL-18.0.0
разгон модуля / 1.63.0 / INTEL-18.0.0
# активируем conda env, в котором мы установили tigmint
conda активировать строительные леса
# убедитесь, что мы вводим все правильные пути
экспорт PATH = / home / brant / project / shared / bin /: / home / brant / project / shared / src / links_v1.8.7: $ PATH
cd $ PBS_O_WORKDIR
сделать -f аркс-make.txt аркс-тигминт \
draft = diglossa.contigs \
читает = штрих-код \
m = 50-30000 o = 3 раза = 1
Механизм сборки вирусов, управляемый каркасами, предложенный сравнением прокапсидов P22, содержащих каркасы и не содержащих каркасов
https://doi.org/10.1016/S0006-3495(99)77479-5 Получить права и контент Резюме
Рекомендуемые статьи
Ссылки на статьи
Расширение строительных лесов и закрытие зазоров
bowtie2-build ALT_04162018_0_2um_scaffold_13.fasta ALT_04162018_0_2um_scaffold_13.fasta bowtie2 -p 6 -X 2000 -x ALT_04162018_0_2um_scaffold_13.fasta -1 ALT_04162018_0_2um.1.fastq.gz -2 ALT_04162018_0_2um.2.fastq.gz | shrinksam -v> ALT_04162018_0_2um_scaffold_13.fasta.mapped.sam Shrinksam - это инструмент для фильтрации исходного файла SAM из Bowtie 2, который будет записывать в файл только отображенные операции чтения и, таким образом, экономить много места для хранения. Shrinksam можно загрузить с Github (https://github.com/bcthomas/shrinksam). Сборка генома квиноа с использованием геномной вариации для управляемого каркаса
Сборка генома на основе информации о гаплотипе
Обнаружение общих отпечатков вариаций с использованием данных
Arabidopsis Сборка генома квиноа
Интеграция сборки с общедоступной картой генетического сцепления
Секвенирование 50 образцов квиноа и варианта, вызывающего
Упорядочивание контигов на основе информации о гаплотипах
Проверка основанных на гаплотипах отношений
Создание каркасов и объединение контигов квиноа
Хромосомы, каркасы и контиги
Хромосомы, каркасы и контиги Верхний уровень