Монтаж строительных лесов – блог ООО Эльба
Категории блога
Популярные записи
19 июля 2018 г.
I-Этап монтажа
Нижний ряд рам устанавливается на опорные пяты или регулируемые опоры. Все элементы должны находиться строго в вертикальном положении! Зависимости от поверхности укладывается деревянное покрытие для амортизации.
II-Этап монтажа
Расстояние между рамами должно составлять 3 метра, тем самым вы сможете соединить их горизонтальными и диагональными связями. По краям устанавливаются рамы ограждения, а следующая должна быть обязательно с лестницей. По необходимости можно добавлять рамы с лестницей для более удобной и быстрой работы.
III-Этап монтажа
Второй ярус рам устанавливается в первый строго вертикально, начиная с рамы с лестницей. Все рамы между собой для жесткости связывают диагональными и горизонтальными связями. Для этого на рамах предусмотрены фиксаторы-флажки.
Для перемещения по лесам необходимо установить ригеля настилов по 2 шт. , на 1 секцию. На ригеля укладываются деревянные настилы размером 1 м х 1 м по 3 шт. на 2 ригеля.
, на 1 секцию. На ригеля укладываются деревянные настилы размером 1 м х 1 м по 3 шт. на 2 ригеля.
IV-Этап монтажа
Важным этапом является, крепление конструкции к стене при помощи кронштейнов. Одна часть кронштейна закрепляется к лесам анкерной частью, другая к стене с помощью дюбель – гвоздей. Шаг крепления происходит через каждые 4 метра.
Элементы анкеровки
с крюком и втулкой
с пробкой
Схема крепления лесов к стене
( Х – место крепления )
Что касается техники безопасности – на фасадной части строительных лесов так же натягивается защитная сетка. Она не позволит различным предметам упасть на тротуар. При сильном ветре она так же защитит Вас от порывов ветра. Придаст эстетический вид вашему объекту.
Что следует учитывать при монтаже конструкций?
Прежде чем сделать свой выбор, стоит:
- Внимательно ознакомиться с характеристикой всех видов строительных лесов и особенностей их установки.

- Далее важно заранее подготовить место для их монтажа (очистить от всего лишнего, выровнять, утрамбовать и сделать водоотводы).
- Для установки конструкций потребуются специальные подъемники.
- Крепежи, горизонтальные площадки, детали ограждений собираются по ходу ведения сборки. Рамные составляющие конструкции требуют особого контроля их установки с помощью отвеса.
- Леса фиксируются во всю высоту здания, для того чтобы обеспечить безопасность строителей и подсобных рабочих. Расположение ровных и прочных настилов должно быть строго горизонтально.
- Передвижение рабочих вверх-вниз обязательно происходит по надежным лестницам.
- Прежде чем демонтировать конструкцию после выполнения работ, следует убрать с ее поверхности все, без исключения, предметы.

Только серьезно и ответственно подходя к выбору и установке строительных лесов, можно обеспечить безопасность и качество производимых работ.
Сборка лесов строительных – монтаж строительных хомутовых лесов, цена аренды и сборки строительных лесов в Астрахани и Краснодаре
| Услуги |
Компания АЛК-КС имеет большой опыт монтажа хомутовых лесов. Установка строительных лесов ответственное дело, от качества сборки зависит прочность всей конструкции. Поэтому сборка лесов строительных должна производиться профессионалами с опытом. За нашими плечами десятки объектов на которых мы занимались сборкой строительных лесов. В их числе установка строительных лесов в Астрахани, монтаж строительных лесов в Краснодаре и Волгограде.
Установка строительных лесов ответственное дело, от качества сборки зависит прочность всей конструкции. Поэтому сборка лесов строительных должна производиться профессионалами с опытом. За нашими плечами десятки объектов на которых мы занимались сборкой строительных лесов. В их числе установка строительных лесов в Астрахани, монтаж строительных лесов в Краснодаре и Волгограде.
Хомутовые строительные леса, используемые нашей компанией для строительства площадок и доступов, представляют собой конструкцию из труб, соединяемых между собой замками. Для последовательного соединения труб используется специальный соединитель, для установки строительных лесов на поверхности — опорные пяты.
Схема расположения труб определяется под особенности объекта: из небольшого числа комплектующих можно возвести конструкцию произвольной формы и произвольного размера. Благодаря этому монтаж хомутовых лесов подойдет для всех видов работ с объектами любой сложности. Цена монтажа строительных лесов не отличается от города. Будь то Волгоград или Краснодар цена монтажа строительных лесов будет одинаковой и рассчитывается из объема и сложности объекта.
Будь то Волгоград или Краснодар цена монтажа строительных лесов будет одинаковой и рассчитывается из объема и сложности объекта.
Хомутовые строительные леса имеют преимущества перед остальными видами строительных лесов, в частности в том, что хомутовые леса можно монтировать в произвольные по форме конструкции, повторяющие пространственную форму строения. Аренда хомутовых лесов так же дешевле чем обычные строительные леса. Мы занимаемся установкой строительных лесов в Краснодаре, при этом вы можете взять в аренду хомутовые леса в Волгограде или Астрахани в наших представительствах. Монтаж строительных лесов, цена зависит от сложности и объемов работ. Благодаря этому заказчики получают удобный и безопасный доступ для проведения работ на высоте, к строениям, имеющим сложную пространственную геометрию, в случаях, когда работа на других видах строительных лесов была бы недостаточно удобной.
Мы занимаемся монтажом и арендой хомутовых лесов, о цене монтажа строительных лесов вы можете узнать в наших представительствах в Астрахани и Краснодаре.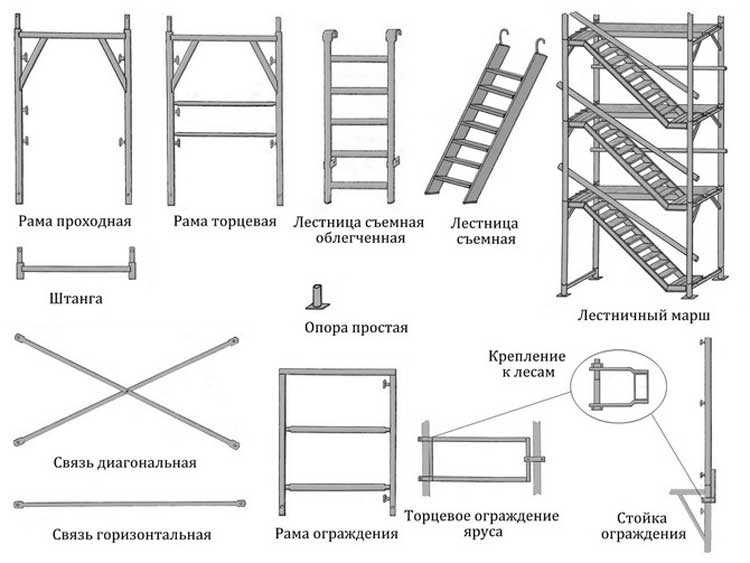 Сборка и установка лесов строительных ответственное дело, доверяйте монтаж только профессионалам.
Сборка и установка лесов строительных ответственное дело, доверяйте монтаж только профессионалам.
Хомутовые строительные леса отлично подходят для самых разных задач: от строительных до судоремонтных работ. Возможно возведение конструкции в точности повторяющей форму строения, поэтому могут применяться на объектах любой сложности.
Малый вес позволяет заниматься сборкой хомутовых строительных лесов быстро, что значительно снижает трудозатраты и расходы на транспортировку. Для удобства у нас есть представительства в Волгограде, Астрахани и Краснодаре – это минимизирует затраты на транспортировку и сборку строительных лесов.
При строительстве Самоподъемных Буровых установок “ Нептун” и “Меркурий“ нашей компанией применялись хомутовые леса, что предельно облегчило строительство площадок для различных уровней труднодоступных и надводных частей строительства.
Нашими специалистами также производится укрытие площадей проведения работ специальным материалом, что позволяет производить работы в любых погодных условиях и предотвращает загрязнение окружающей среды.
Возврат к списку
всесторонний обзор методов каркаса в сборке генома | Брифинги по биоинформатике
Статья журнала
Получить доступ
Цзюньвэй Луо,
Цзюньвэй Луо
Ищите другие работы этого автора на:
Оксфордский академический
пабмед
Google Scholar
Явэй Вэй,
Явэй Вэй
Ищите другие работы этого автора на:
Оксфордский академический
пабмед
Google Scholar
Менгна Лю,
Менгна Лю
Ищите другие работы этого автора на:
Оксфордский академический
пабмед
Google Scholar
Чжэнцзян Ву,
Чжэнцзян Ву
Ищите другие работы этого автора на:
Оксфордский академический
пабмед
Google Scholar
Сяоянь Лю,
Сяоянь Лю
Ищите другие работы этого автора на:
Оксфордский академический
пабмед
Google Scholar
Хуйминь Луо,
Хуимин Луо
Ищите другие работы этого автора на:
Оксфордский академический
пабмед
Google Scholar
Чаокун Ян
Чаокун Ян
Ищите другие работы этого автора на:
Оксфордский академический
пабмед
Google Scholar
Брифинги по биоинформатике , том 22, выпуск 5, сентябрь 2021 г. , bbab033, https://doi.org/10.1093/bib/bbab033
, bbab033, https://doi.org/10.1093/bib/bbab033
Опубликовано:
25 февраля 2021 г.
История статьи
9 0002- Содержание статьи
- Рисунки и таблицы
- видео
- Аудио
- Дополнительные данные
Цитировать
Cite
Junwei Luo, Yawei Wei, Mengna Lyu, Zhengjiang Wu, Xiaoyan Liu, Huimin Luo, Chaokun Yan, Всесторонний обзор методов каркаса при сборке генома,  , bbab033, https://doi.org/10.1093/bib/bbab033
, bbab033, https://doi.org/10.1093/bib/bbab033
Выберите формат Выберите format.ris (Mendeley, Papers, Zotero).enw (EndNote).bibtex (BibTex).txt (Medlars, RefWorks)
Закрыть
Разрешения
- Электронная почта
- Твиттер
- Фейсбук
- Подробнее
Фильтр поиска панели навигации Брифинги по биоинформатикеЭтот выпускБиоинформатика и вычислительная биологияКнигиЖурналыOxford Academic Термин поиска мобильного микросайта
Закрыть
Фильтр поиска панели навигации Брифинги по биоинформатикеЭтот выпускБиоинформатика и вычислительная биологияКнигиЖурналыOxford Academic Термин поиска на микросайте
Advanced Search
Abstract
В области сборки генома методы каркаса позволяют получить более полный и непрерывный эталонный геном, что является краеугольным камнем геномных исследований.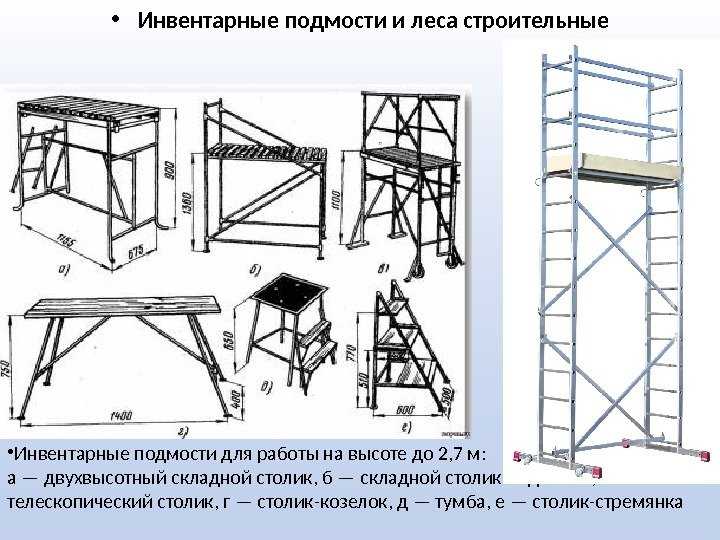

Сборка генома, создание каркасов, выравнивание, секвенирование на большие расстояния
© Автор(ы), 2021 г. Опубликовано Oxford University Press. Все права защищены. Для получения разрешений обращайтесь по электронной почте: [email protected]
Раздел проблем:
Проверка метода
В настоящее время у вас нет доступа к этой статье.
Скачать все слайды
Войти
Получить помощь с доступом
Получить помощь с доступом
Доступ для учреждений
Доступ к контенту в Oxford Academic часто предоставляется посредством институциональных подписок и покупок. Если вы являетесь членом учреждения с активной учетной записью, вы можете получить доступ к контенту одним из следующих способов:
Доступ на основе IP
Как правило, доступ предоставляется через институциональную сеть к диапазону IP-адресов. Эта аутентификация происходит автоматически, и невозможно выйти из учетной записи с IP-аутентификацией.
Эта аутентификация происходит автоматически, и невозможно выйти из учетной записи с IP-аутентификацией.
Войдите через свое учреждение
Выберите этот вариант, чтобы получить удаленный доступ за пределами вашего учреждения. Технология Shibboleth/Open Athens используется для обеспечения единого входа между веб-сайтом вашего учебного заведения и Oxford Academic.
- Щелкните Войти через свое учреждение.
- Выберите свое учреждение из предоставленного списка, после чего вы перейдете на веб-сайт вашего учреждения для входа в систему.
- Находясь на сайте учреждения, используйте учетные данные, предоставленные вашим учреждением. Не используйте личную учетную запись Oxford Academic.
- После успешного входа вы вернетесь в Oxford Academic.
Если вашего учреждения нет в списке или вы не можете войти на веб-сайт своего учреждения, обратитесь к своему библиотекарю или администратору.
Вход с помощью читательского билета
Введите номер своего читательского билета, чтобы войти в систему. Если вы не можете войти в систему, обратитесь к своему библиотекарю.
Члены общества
Доступ члена общества к журналу достигается одним из следующих способов:
Войти через сайт сообщества
Многие общества предлагают единый вход между веб-сайтом общества и Oxford Academic. Если вы видите «Войти через сайт сообщества» на панели входа в журнале:
- Щелкните Войти через сайт сообщества.
- При посещении сайта общества используйте учетные данные, предоставленные этим обществом. Не используйте личную учетную запись Oxford Academic.
- После успешного входа вы вернетесь в Oxford Academic.
Если у вас нет учетной записи сообщества или вы забыли свое имя пользователя или пароль, обратитесь в свое общество.
Вход через личный кабинет
Некоторые общества используют личные аккаунты Oxford Academic для предоставления доступа своим членам. Смотри ниже.
Личный кабинет
Личную учетную запись можно использовать для получения оповещений по электронной почте, сохранения результатов поиска, покупки контента и активации подписок.
Некоторые общества используют личные аккаунты Oxford Academic для предоставления доступа своим членам.
Просмотр учетных записей, вошедших в систему
Щелкните значок учетной записи в правом верхнем углу, чтобы:
- Просмотр вашей личной учетной записи, в которой выполнен вход, и доступ к функциям управления учетной записью.
- Просмотр институциональных учетных записей, предоставляющих доступ.
Выполнен вход, но нет доступа к содержимому
Oxford Academic предлагает широкий ассортимент продукции. Подписка учреждения может не распространяться на контент, к которому вы пытаетесь получить доступ. Если вы считаете, что у вас должен быть доступ к этому контенту, обратитесь к своему библиотекарю.
Подписка учреждения может не распространяться на контент, к которому вы пытаетесь получить доступ. Если вы считаете, что у вас должен быть доступ к этому контенту, обратитесь к своему библиотекарю.
Ведение счетов организаций
Для библиотекарей и администраторов ваша личная учетная запись также предоставляет доступ к управлению институциональной учетной записью. Здесь вы найдете параметры для просмотра и активации подписок, управления институциональными настройками и параметрами доступа, доступа к статистике использования и т. д.
Покупка
Стоимость подписки и заказ этого журнала
Варианты покупки книг и журналов в Oxford Academic
Кратковременный доступ
Чтобы приобрести краткосрочный доступ, пожалуйста, войдите в свой личный аккаунт выше.
У вас еще нет личного кабинета? регистр
Всесторонний обзор методов каркаса при сборке генома — круглосуточный доступ
ЕВРО €30,00
22 фунта стерлингов
39 долларов США.
Реклама
Цитаты
Альтметрика
Дополнительная информация о метриках
Оповещения по электронной почте
Оповещение об активности статьи
Предварительные уведомления о статьях
Оповещение о новой проблеме
Оповещение о текущей проблеме
Получайте эксклюзивные предложения и обновления от Oxford Academic
Ссылки на статьи по телефону
Последний
Самые читаемые
Самые цитируемые
Оценка алгоритмов маркировки РНКсек одиночных клеток с использованием наборов данных о раке
Всестороннее исследование статистических подходов и подходов машинного обучения для прогнозирования сложных заболеваний человека на основе геномных вариантов
Сравнение матриц сходства для анализа основных компонентов для оценки стратификации населения в секвенированных наборах генетических данных
Вывод сетей регуляции генов из данных об экспрессии генов в одной клетке с помощью глубокого многопрофильного контрастного обучения
Открытие слитых кольцевых РНК при лейкемии с перестройками KMT2A::AFF1 с помощью нового программного обеспечения CircFusion
Реклама
ntJoin: быстрое и легкое построение каркаса под управлением сборки с использованием графов минимизации
- Список журналов
- Биоинформатика
- PMC7320612
Биоинформатика. 2020 15 июня; 36 (12): 3885–3887.
2020 15 июня; 36 (12): 3885–3887.
Опубликовано онлайн 2020 апреля 20. DOI: 10.1093/Bioinformatics/BTAA253
, , , , и
Alfonso Valenci Материалы
Резюме
Возможность генерировать высококачественные последовательности генома является краеугольным камнем современных биологических исследований. Даже с недавними достижениями в технологиях секвенирования многие сборки генома все еще не достигают эталонного уровня. Здесь мы представляем ntJoin, инструмент, который использует структурную синтению между черновой сборкой и ссылочной последовательностью(ями) для сопряжения и исправления первой по отношению к последней. Вместо выравнивания ntJoin использует упрощенный подход к сопоставлению, основанный на структуре данных графа, созданной из упорядоченных эскизов минимизатора. Этот инструмент можно использовать в различных приложениях, в том числе для улучшения черновой сборки с помощью генома эталонного уровня, сборки для короткого чтения с черновой сборкой для длинного чтения и черновой сборки для сборки из близкородственного вида. При построении сборки короткого считывания человека с использованием эталонного генома человека или сборки длинного считывания ntJoin улучшает длину NGA50 в 23 и 13 раз соответственно при длине менее 13 м и использовании менее 11 ГБ ОЗУ. По сравнению с существующими скаффолдерами, основанными на ссылках, ntJoin создает очень непрерывные сборки быстрее и использует меньше памяти.
При построении сборки короткого считывания человека с использованием эталонного генома человека или сборки длинного считывания ntJoin улучшает длину NGA50 в 23 и 13 раз соответственно при длине менее 13 м и использовании менее 11 ГБ ОЗУ. По сравнению с существующими скаффолдерами, основанными на ссылках, ntJoin создает очень непрерывные сборки быстрее и использует меньше памяти.
Доступность и реализация
ntJoin написан на C++ и Python и находится в свободном доступе на https://github.com/bcgsc/ntjoin.
Дополнительная информация
Дополнительные данные доступны по адресу Bioinformatics онлайн.
Создание сильно непрерывных сборок позволяет проводить важные последующие исследования, такие как исследования генетической ассоциации и анализ цис--регуляторных элементов (Rice and Green, 2019). Однако, несмотря на то, что продвижение данных секвенирования отдельных молекул, таких как связанные чтения и длинные чтения, показало большие перспективы в улучшении de novo качество сборки генома (Shafin et al. , 2019; Weisenfeld et al. , 2017), большинство черновых сборок все еще не достигают полноты хромосомного масштаба.
, 2019; Weisenfeld et al. , 2017), большинство черновых сборок все еще не достигают полноты хромосомного масштаба.
Для некоторых черновиков геномов могут быть доступны более смежные сборки для разных особей того же вида или даже близкородственных видов. В этом случае синхронизация последовательности между сборками может быть использована для построения лесов, управляемых сборкой. Например, в то время как сборки с длительным чтением могут генерировать непрерывные черновики геномов, высокая частота ошибок чтения отрицательно влияет на базовое качество, затрудняя аннотацию генов (Watson and Warr, 2019).). Полировка с использованием коротких считываний часто используется для повышения точности сборки в паре оснований (Rice and Green, 2019; Warren et al. , 2019; Watson and Warr, 2019). Альтернативный подход к этому шагу полировки состоит в том, чтобы собрать короткие риды отдельно и построить сборку коротких ридов, используя сборку длинных ридов, создавая сборку наравне с непрерывностью и структурой сборки длинных ридов и точностью пары оснований сборка короткого чтения.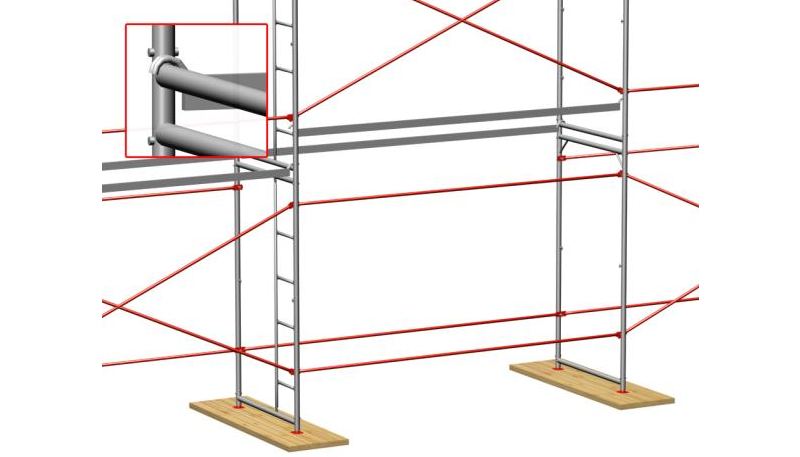
Существующие опорные леса, такие как Ragout (Колмогоров и др. , 2018) и Ragoo (Alonge et al. , 2019) полагаются на выравнивание черновой сборки с эталонной сборкой; Ragout использует Progressive Cactus (Armstrong et al. , 2019) для больших геномов, в то время как Ragoo использует для этой задачи minimap2 (Li, 2018). Использование набросков минимизатора в таких инструментах, как minimap2, очень эффективно для компактного представления последовательностей генома. Вместо того, чтобы сохранять каждое слово размера k (k-мер) из входных последовательностей, сохраняется только выбранный набор k-меров или хэш-значений («минимизаторы»), что значительно снижает вычислительные затраты на хранение данных последовательности и манипулирование ими (Робертс и др. , 2004).
Здесь мы представляем ntJoin, скаффолдер, управляемый сборкой, который использует упрощенную стратегию сопоставления без выравнивания вместо выравнивания для быстрого сопряжения целевой сборки с использованием одной или нескольких ссылок.
Учитывая входную целевую и эталонную последовательности в формате fasta, ntJoin сначала создает упорядоченный эскиз минимизатора для каждого из предоставленных наборов последовательностей, как описано ранее (Roberts et al. , 2004) (дополнительные рисунки S1 и С2). Затем ntJoin использует упорядоченные эскизы минимизатора из каждого входа для построения единого неориентированного графа, который облегчает легкое сопоставление между ними. В этом графе каждый узел является минимайзером, а ребра создаются между минимизаторами, соседними хотя бы в одном из упорядоченных эскизов. Вес ребра — это мера, которая используется для того, чтобы сделать больший акцент на соединениях в определенных входных сборках. Веса ребер представляют собой сумму заданных пользователем весов каждого входа, который поддерживает это ребро.
Затем граф подвергается серии шагов фильтрации. Во-первых, применяется глобальный порог веса ребра. Затем идентифицируются узлы ветвления (узлы со степенью > 2), и инцидентные ребра фильтруются с возрастающим пороговым значением веса ребра до тех пор, пока степень этого узла не упадет до <3. Как правило, вес эталона выше, чем вес целевой сборки, что приводит к тому, что эти ребра имеют приоритет, и в результате целевая сборка подгоняется к эталонной структуре. Фильтрация инцидентных ребер узлов ветвления приводит к тому, что граф представляет собой набор компонент связности, каждая из которых представляет собой линейный путь узлов-минимизаторов. Последовательности минимизаторов в линейных путях затем переводятся в упорядоченные и ориентированные непрерывные пути, которые описывают окончательные выходные каркасы (дополнительные рисунки S1 – S3). Этот основанный на графе метод позволяет алгоритму выполнять исправление неправильной сборки в дополнение к формированию входных контигов на основе входной эталонной сборки, поскольку контиги могут быть нарушены при предполагаемых ошибках сборки в режиме по умолчанию. Если пользователь не хочет, чтобы входные контиги обрезались при подгонке эталонной последовательности, можно указать параметр «no_cut=True».
Как правило, вес эталона выше, чем вес целевой сборки, что приводит к тому, что эти ребра имеют приоритет, и в результате целевая сборка подгоняется к эталонной структуре. Фильтрация инцидентных ребер узлов ветвления приводит к тому, что граф представляет собой набор компонент связности, каждая из которых представляет собой линейный путь узлов-минимизаторов. Последовательности минимизаторов в линейных путях затем переводятся в упорядоченные и ориентированные непрерывные пути, которые описывают окончательные выходные каркасы (дополнительные рисунки S1 – S3). Этот основанный на графе метод позволяет алгоритму выполнять исправление неправильной сборки в дополнение к формированию входных контигов на основе входной эталонной сборки, поскольку контиги могут быть нарушены при предполагаемых ошибках сборки в режиме по умолчанию. Если пользователь не хочет, чтобы входные контиги обрезались при подгонке эталонной последовательности, можно указать параметр «no_cut=True».
Окончательная целевая сборка в формате fasta является основным результатом работы ntJoin. Кроме того, сведения о том, как была создана целевая сборка, включая ориентацию, порядок и размеры зазоров, описываются в выходном файле «пути» и, необязательно, в файле agp (опция «agp = True»). Наконец, граф минимизатора выводится в «точечном» формате, который дает все узлы и ребра в графе, а также веса ребер, указывающие уровень поддержки сборки.
Кроме того, сведения о том, как была создана целевая сборка, включая ориентацию, порядок и размеры зазоров, описываются в выходном файле «пути» и, необязательно, в файле agp (опция «agp = True»). Наконец, граф минимизатора выводится в «точечном» формате, который дает все узлы и ребра в графе, а также веса ребер, указывающие уровень поддержки сборки.
Каждая сборка оценивалась на непрерывность и правильность с помощью QUAST (v5.0.2; –scaffold-gap-max-size 100 000 –large) (Михеенко и др. , 2018). Эта настройка параметра зазора каркаса приводит к тому, что несоответствия размера зазора более 100 КБ классифицируются как «обширные неправильные сборки».
Подробные методы доступны в Интернете.
Сначала мы протестировали ntJoin с использованием различных сборок образцов Caenorhabditis elegans и Homo sapiens (дополнительные таблицы S1 и S2). По сравнению с Ragout и Ragoo, ntJoin обычно создает сборки с большей длиной NGA50 (длина, которая захватывает не менее 50% генома, используя длины выравнивания с эталоном вместо длин контигов) и сопоставимые или меньшие неправильные сборки (; дополнительные рисунки S4 –S10; Дополнительные таблицы S3–S11). Примечательно, что ntJoin улучшает сборки с начальной смежностью в диапазоне килобаз, чтобы достичь масштаба в мегабазах (NGA50 увеличивается с 26,9kb до 2,3 Мбит/с и 19,8 kb до 50,3 Мбит/с для сборок с коротким чтением C.elegans и H.sapiens соответственно, дополнительные рисунки S4 и S6, при этом количество неправильных сборок сократилось более чем на треть (33,5 и 61,5% , соответственно). Это подчеркивает потенциал ntJoin в улучшении фрагментированных черновых сборок. По сравнению с Ragout, ntJoin достиг значений NGA50 в 1,1-2 раза выше для протестированных сборок ABySS с коротким считыванием, хотя Ragout действительно поднял сборку Shasta с длинным считыванием до 1,2-кратного увеличения NGA50 (; Дополнительные рисунки S4 и S6) ( Джекман и др. , 2017; Шафин и др. , 2019). Однако выравнивание Progressive Cactus, необходимое для Ragout, было очень затратным с точки зрения вычислений, выполнялось более четырех дней для всех запусков вручную и использовало более 115 ГБ ОЗУ по сравнению с запусками ntJoin для людей, которые завершились менее чем за 13 минут и использовали <11 ГБ памяти.
Примечательно, что ntJoin улучшает сборки с начальной смежностью в диапазоне килобаз, чтобы достичь масштаба в мегабазах (NGA50 увеличивается с 26,9kb до 2,3 Мбит/с и 19,8 kb до 50,3 Мбит/с для сборок с коротким чтением C.elegans и H.sapiens соответственно, дополнительные рисунки S4 и S6, при этом количество неправильных сборок сократилось более чем на треть (33,5 и 61,5% , соответственно). Это подчеркивает потенциал ntJoin в улучшении фрагментированных черновых сборок. По сравнению с Ragout, ntJoin достиг значений NGA50 в 1,1-2 раза выше для протестированных сборок ABySS с коротким считыванием, хотя Ragout действительно поднял сборку Shasta с длинным считыванием до 1,2-кратного увеличения NGA50 (; Дополнительные рисунки S4 и S6) ( Джекман и др. , 2017; Шафин и др. , 2019). Однако выравнивание Progressive Cactus, необходимое для Ragout, было очень затратным с точки зрения вычислений, выполнялось более четырех дней для всех запусков вручную и использовало более 115 ГБ ОЗУ по сравнению с запусками ntJoin для людей, которые завершились менее чем за 13 минут и использовали <11 ГБ памяти. БАРАН. ntJoin также был быстрее Ragoo во всех тестах: от 1,4 раза быстрее для сборки Shasta до 35,8 раза быстрее для более фрагментированной сборки H.sapiens ABySS (дополнительные рисунки S4 и S6). ntПрисоединиться к местам 86,3–99,3% входной сборки в лесах, пропорция, которая очень похожа как на Ragout, так и на Ragoo. Последовательности не могут быть помещены в каркасы, если они слишком короткие по сравнению с установленным пользователем размером окна или если выбранная ссылка слишком расходится. В то время как Ragoo использует постоянный размер промежутка между соединяемыми контигами (по умолчанию 100 bp), ntJoin и Ragout оценивают размеры промежутка на основе эталона, о чем свидетельствует общий размер промежутка в каркасных сборках Ragoo, который значительно меньше, чем у ntJoin и Ragout (дополнительная информация). Таблицы S4–S15).
БАРАН. ntJoin также был быстрее Ragoo во всех тестах: от 1,4 раза быстрее для сборки Shasta до 35,8 раза быстрее для более фрагментированной сборки H.sapiens ABySS (дополнительные рисунки S4 и S6). ntПрисоединиться к местам 86,3–99,3% входной сборки в лесах, пропорция, которая очень похожа как на Ragout, так и на Ragoo. Последовательности не могут быть помещены в каркасы, если они слишком короткие по сравнению с установленным пользователем размером окна или если выбранная ссылка слишком расходится. В то время как Ragoo использует постоянный размер промежутка между соединяемыми контигами (по умолчанию 100 bp), ntJoin и Ragout оценивают размеры промежутка на основе эталона, о чем свидетельствует общий размер промежутка в каркасных сборках Ragoo, который значительно меньше, чем у ntJoin и Ragout (дополнительная информация). Таблицы S4–S15).
Открыть в отдельном окне
Сравнение ( a ) непрерывности, правильности и ( b ) результатов бенчмаркинга ntJoin (оранжевый), Ragoo (синий) и Ragout (зеленый) на различных H. sapiens (NA12878) сборки в линейном (а) и логарифмическом масштабе (б). Эталонные геномы представляют собой эталонный геном человека («Ref») и отполированную с помощью ntEdit сборку Shasta («Shasta»). Улучшаемые целевые сборки — это сборка NA12878 ABySS, созданная с использованием данных MPET («ABySS»), и сборка Shasta, отполированная с помощью ntEdit («Shasta»). «Базовая» статистика показана для соответствующих целевых сборок до возведения лесов на каждой панели (а)
sapiens (NA12878) сборки в линейном (а) и логарифмическом масштабе (б). Эталонные геномы представляют собой эталонный геном человека («Ref») и отполированную с помощью ntEdit сборку Shasta («Shasta»). Улучшаемые целевые сборки — это сборка NA12878 ABySS, созданная с использованием данных MPET («ABySS»), и сборка Shasta, отполированная с помощью ntEdit («Shasta»). «Базовая» статистика показана для соответствующих целевых сборок до возведения лесов на каждой панели (а)
ntJoin также может улучшить черновики сборок, если для одного и того же вида доступны смежные сборки. Это типичный вариант использования в проекте, использующем несколько платформ секвенирования для гибридной сборки. В 2019 году сборка ABySS с коротким чтением была построена с использованием сборки Shasta с длинным чтением (Shafin et al. , 2019). Сохраняя соединения, уникальные для длинных и коротких последовательностей чтения, ntJoin достигает NGA50 выше, чем базовая сборка Shasta. ntJoin предоставляет альтернативный конвейер сборки, в котором структура сборки с длительным чтением сообщает о размещении последовательностей сборок с коротким чтением, исключая необходимость полировки сборки с длительным чтением с помощью коротких операций чтения.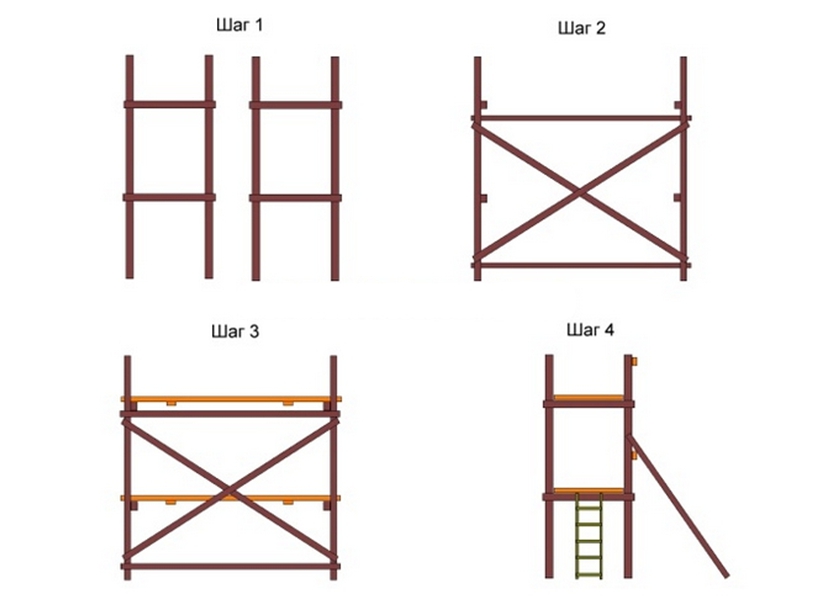 Этот подход может создавать сборки с высокой непрерывностью и базовой точностью, что особенно важно для аннотации нижестоящего генома (дополнительные таблицы S12–S15). Согласно этим данным, ни Ragout, ни Ragoo не дают сборок с одинаково высокой длиной NGA50, и обе требуют больше времени и памяти по сравнению с ntJoin.
Этот подход может создавать сборки с высокой непрерывностью и базовой точностью, что особенно важно для аннотации нижестоящего генома (дополнительные таблицы S12–S15). Согласно этим данным, ни Ragout, ни Ragoo не дают сборок с одинаково высокой длиной NGA50, и обе требуют больше времени и памяти по сравнению с ntJoin.
Подход ntJoin также распространяется на сборки лесов разных видов, как было продемонстрировано созданием лесов сборок морских и гавиальных крокодилов с использованием генома американского аллигатора в качестве эталона (дополнительные таблицы S16 и S17). В наших тестах длина NG50 крокодиловых сборок увеличилась до 14,44 и 12,92 Мбн для морских и гавиальных крокодилов (исходный уровень NG50 = 0,14 и 0,07 Мбп) с соответствующим увеличением полноты генов BUSCO (Simão et al. , 2015). 2,6 и 90,5% соответственно. Это показывает, что ntJoin все еще может использовать синтению между этими целевыми и эталонными сборками, несмотря на то, что виды разошлись около 80 миллионов лет назад (Delsuc et al. , 2018).
, 2018).
Любому инструменту сборки, основанному на эталонах, присуще некоторое смещение эталона, и пользователь должен учитывать это при разработке своего эксперимента, в том числе при выборе эталонной сборки. Здесь мы демонстрируем полезность ntJoin для подгонки входной целевой сборки к структуре эталона, что исправляет неправильные сборки, но также потенциально ломает некоторые большие структурные варианты. Точно так же инструменты компаратора были запущены в режимах, которые также могут обрезать входные контиги. Хотя этот режим не нарушит все структурные изменения (дополнительная таблица S18), чтобы избежать разрыва/обрезания входных контигов, можно указать параметр ntJoin no_cut = True, который предотвращает стирание любых существующих структурных изменений в целевой сборке (дополнительный рис. С11).
В заключение следует отметить, что ntJoin быстро и с небольшим объемом памяти выполняет минимизацию построения каркасов на основе графов, сохраняя при этом смежность на уровне хромосом. Как продемонстрировано, это гибкий, не требующий выравнивания инструмент построения каркаса, который можно использовать в ряде различных приложений, включая гибридную сборку и исследования популяционной геномики.
Как продемонстрировано, это гибкий, не требующий выравнивания инструмент построения каркаса, который можно использовать в ряде различных приложений, включая гибридную сборку и исследования популяционной геномики.
Эта работа была поддержана Genome BC и Genome Canada [243FOR, 281ANV]; и Национальных институтов здравоохранения [2R01HG007182-04A1]. Ответственность за содержание этой статьи несут исключительно авторы, и она не обязательно отражает официальную точку зрения Национальных институтов здравоохранения или других финансирующих организаций.
Конфликт интересов : не объявлено.
btaa253_Supplementary_Data
Щелкните здесь, чтобы просмотреть файл с дополнительными данными. (1.3M, pdf)
- Аугле М. и другие. (2019) RaGOO: быстрое и точное построение черновиков геномов с помощью справочных материалов. Genome Biol., 20, 17. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Армстронг Дж.
и другие. (2019) Прогрессивное выравнивание с Cactus: выравниватель нескольких геномов для эпохи тысячи геномов. биоРксив , 730531.
- Делсук Ф. и другие. (2018) Филогеномная основа и временная шкала для сравнительных исследований оболочников. BMC Biol., 16, 39. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Джекман С.Д. и другие. (2017) ABySS 2.0: ресурсоэффективная сборка больших геномов с использованием фильтра Блума. Genome Res., 27, 768–777. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Колмогоров М. и другие. (2018) Хромосомная сборка больших и сложных геномов с использованием нескольких ссылок. Genome Res., 28, 1720–1732. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Ли Х. (2018) Minimap2: попарное выравнивание нуклеотидных последовательностей. Биоинформатика, 34, 3094–3100. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Михеенко А. и другие. (2018) Универсальная оценка сборки генома с помощью QUAST-LG. Биоинформатика, 34, i142–i150. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Райс Э.С., Грин Р.Э.
 биоРксив , 730531.
биоРксив , 730531.